

Modulation of viral immunoinflammatory responses with cytokine DNA administered by different routes
- PMID: 9621011
- PMCID: PMC110202
- DOI: 10.1128/JVI.72.7.5545-5551.1998
Modulation of viral immunoinflammatory responses with cytokine DNA administered by different routes
Abstract
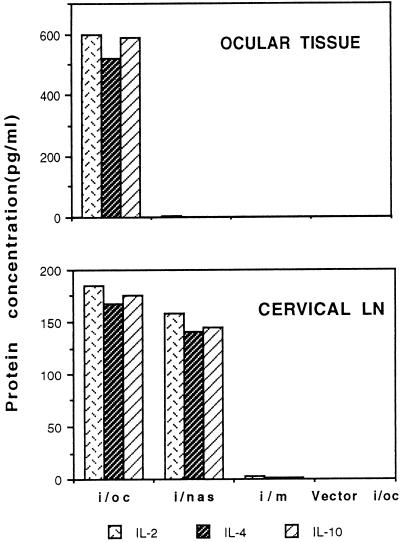
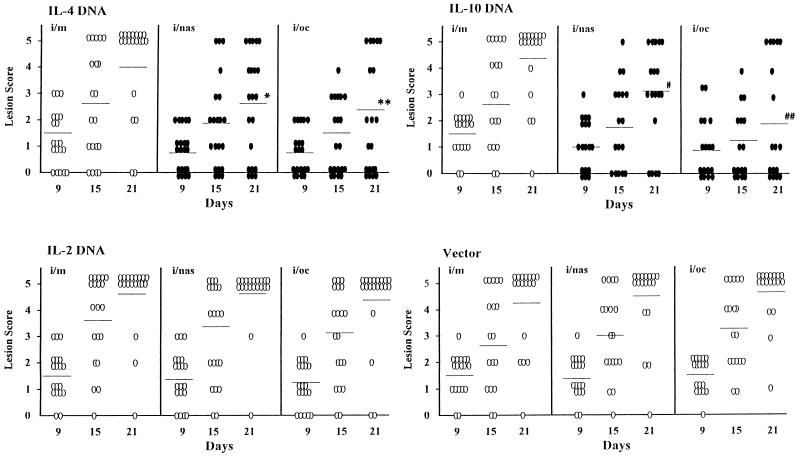
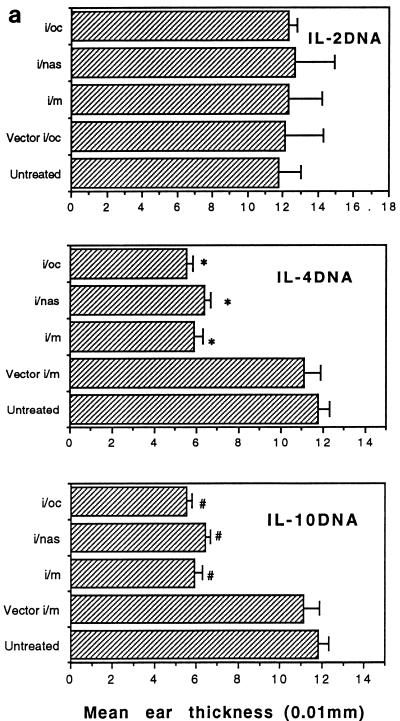
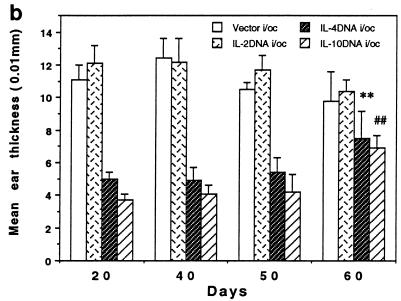
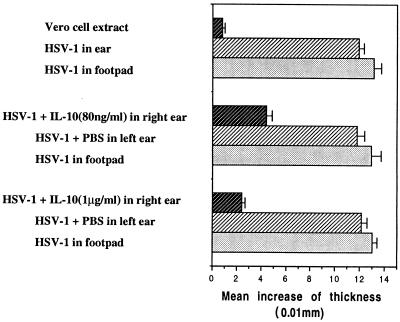
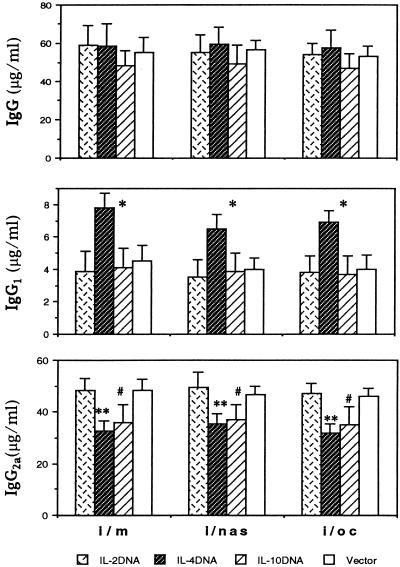
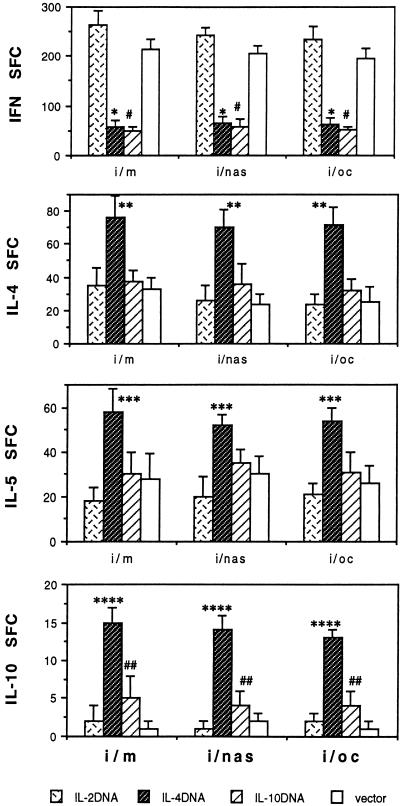
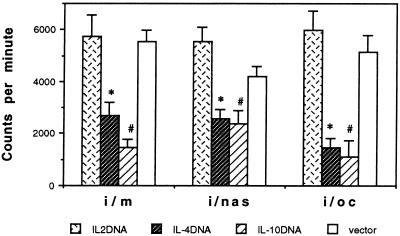
The efficacy of plasmid DNA encoding cytokine administered by different routes, systemic or surface exposure, was evaluated and compared for their modulating effects on subsequent lesions caused by infection with herpes simplex virus (HSV). Systemic or topical administration of both interleukin-4 (IL-4) and IL-10 DNA but not IL-2 DNA caused a long-lasting suppression of HSV-specific delayed-type hypersensitivity response. IL-4 or IL-10 DNA preadministration also modulated the expression of immunoinflammatory lesions associated with corneal infection of HSV. Suppression of ocular lesions required that the DNA be administered to the nasal mucosa or ocular surfaces and was not evident after intramuscular administration. The modulating effect of IL-10 DNA was most evident after topical ocular administration, whereas the effects of IL-4 DNA given by both routes appeared to be equal. Preexposure of IL-4 DNA, but not IL-10 DNA, resulted in a significant change in Th subset balance following HSV infection. Our results indicate that the modulating effect of IL-4 or IL-10 DNA may proceed by different mechanisms. Furthermore, our results suggest that surface administration of cytokine DNA is a convenient means of modulating immunoinflammatory lesions.
Figures
References
-
- Chow Y-H, Chiang B-L, Lee Y-L, Chi W-K, Liu W-C, Chen Y-T, Tao M-H. Development of Th1 and Th2 populations and the nature of immune responses to hepatitis B virus DNA vaccines can be modulated by codelivery of various cytokine genes. J Immunol. 1998;160:1320–1329. - PubMed
-
- Daheshia M, Kuklin N, Kanangat S, Manickan E, Rouse B T. Suppression of ongoing ocular inflammatory disease by topical administration of plasmid DNA encoding IL-10. J Immunol. 1997;159:1945–1952. - PubMed
-
- Daheshia, M., N. Kuklin, E. Manickan, S. Chun, and B. T. Rouse. Immune induction and modulation by topical ocular administration of plasmid DNA encoding antigens and cytokines. Vaccine, in press. - PubMed
-
- Ertl H C J, Xiang Z. Novel DNA vaccine approaches. J Immunol. 1996;156:3579–3582. - PubMed
-
- Groux H, O’Garra A, Bigler M, Rouleau M, Antonenko S, de Vries J E, Roncarolo M G. A CD4+ T cell subset inhibits antigen-specific T cell responses and prevents colitis. Nature. 1997;389:737–742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
