

Genes required for replication of the 15.5-kilobase RNA genome of a plant closterovirus
- PMID: 9621048
- PMCID: PMC110390
- DOI: 10.1128/JVI.72.7.5870-5876.1998
Genes required for replication of the 15.5-kilobase RNA genome of a plant closterovirus
Abstract
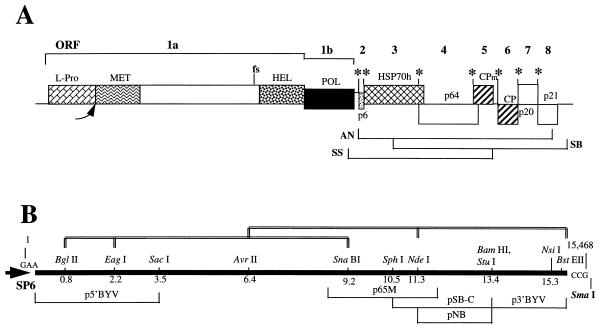
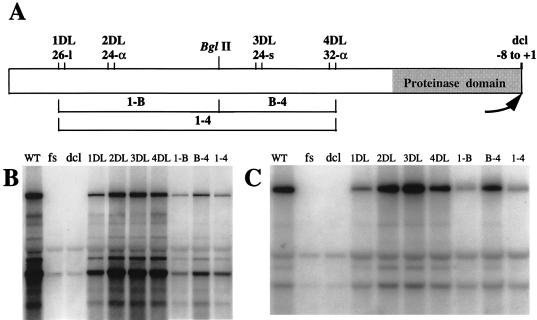
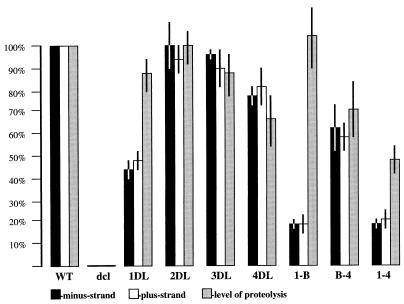
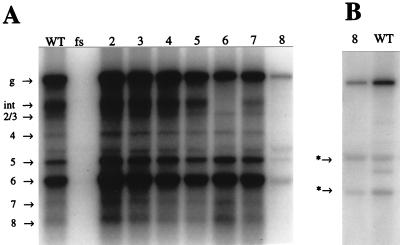
A full-length cDNA clone of beet yellows closterovirus (BYV) was engineered and used to map functions involved in the replication of the viral RNA genome and subgenomic RNA formation. Among 10 open reading frames (ORFs) present in BYV, ORFs 1a and 1b suffice for RNA replication and transcription. The proteins encoded in these ORFs harbor putative methyltransferase, RNA helicase, and RNA polymerase domains common to Sindbis virus-like viruses and a large interdomain region that is unique to closteroviruses. The papain-like leader proteinase (L-Pro) encoded in the 5'-proximal region of ORF 1a was found to have a dual function in genome amplification. First, the autocatalytic cleavage between L-Pro and the remainder of the ORF 1a product was essential for replication of RNA. Second, an additional L-Pro function that was separable from proteolytic activity was required for efficient RNA accumulation. The deletion of a large, approximately 5.6-kb, 3'-terminal region coding for a 6-kDa hydrophobic protein, an HSP70 homolog, a 64-kDa protein, minor and major capsid proteins, a 20-kDa protein, and a 21-kDa protein (p21) resulted in replication-competent RNA. However, examination of mutants with replacements of start codons in each of these seven 3'-terminal ORFs revealed that p21 functions as an enhancer of genome amplification. The intriguing analogies between the genome organization and replicational requirements of plant closteroviruses and animal coronavirus-like viruses are discussed.
Figures

Similar articles
-
Structure and Expression of Large (+)RNA Genomes of Viruses of Higher Eukaryotes.Biochemistry (Mosc). 2021 Mar;86(3):248-261. doi: 10.1134/S0006297921030020. Biochemistry (Mosc). 2021. PMID: 33838627 Free PMC article. Review.
-
Beet yellows closterovirus: complete genome structure and identification of a leader papain-like thiol protease.Virology. 1994 Jan;198(1):311-24. doi: 10.1006/viro.1994.1034. Virology. 1994. PMID: 8259666
-
Complete sequence of the citrus tristeza virus RNA genome.Virology. 1995 Apr 20;208(2):511-20. doi: 10.1006/viro.1995.1182. Virology. 1995. PMID: 7747424
-
Organization of the 3'-terminal half of beet yellow stunt virus genome and implications for the evolution of closteroviruses.Virology. 1996 Jul 1;221(1):199-207. doi: 10.1006/viro.1996.0366. Virology. 1996. PMID: 8661428
-
Citrus tristeza virus: a pathogen that changed the course of the citrus industry.Mol Plant Pathol. 2008 Mar;9(2):251-68. doi: 10.1111/j.1364-3703.2007.00455.x. Mol Plant Pathol. 2008. PMID: 18705856 Free PMC article. Review.
Cited by
-
Hypovirus papain-like protease p29 functions in trans to enhance viral double-stranded RNA accumulation and vertical transmission.J Virol. 2003 Nov;77(21):11697-707. doi: 10.1128/jvi.77.21.11697-11707.2003. J Virol. 2003. PMID: 14557655 Free PMC article.
-
Grapevine leafroll-associated virus 3.Front Microbiol. 2013 Apr 16;4:82. doi: 10.3389/fmicb.2013.00082. eCollection 2013. Front Microbiol. 2013. PMID: 23596440 Free PMC article.
-
The p23 protein of citrus tristeza virus controls asymmetrical RNA accumulation.J Virol. 2002 Jan;76(2):473-83. doi: 10.1128/jvi.76.2.473-483.2002. J Virol. 2002. PMID: 11752137 Free PMC article.
-
Beet mosaic virus expression of a betalain transcription factor allows visual virus tracking in Beta vulgaris.Mol Plant Pathol. 2023 Oct;24(10):1319-1329. doi: 10.1111/mpp.13372. Epub 2023 Jul 6. Mol Plant Pathol. 2023. PMID: 37410356 Free PMC article.
-
Structure and Expression of Large (+)RNA Genomes of Viruses of Higher Eukaryotes.Biochemistry (Mosc). 2021 Mar;86(3):248-261. doi: 10.1134/S0006297921030020. Biochemistry (Mosc). 2021. PMID: 33838627 Free PMC article. Review.
References
-
- Agranovsky A A, Boyko V P, Karasev A V, Koonin E V, Dolja V V. Putative 65kDa protein of beet yellows closterovirus is a homologue of HSP70 heat shock proteins. J Mol Biol. 1991;217:603–610. - PubMed
-
- Agranovsky A A, Boyko V P, Karasev A V, Lunina N A, Koonin E V, Dolja V V. Nucleotide sequence of the 3′-terminal half of beet yellows closterovirus RNA genome: unique arrangement of eight virus genes. J Gen Virol. 1991;72:15–23. - PubMed
-
- Agranovsky A A, Koonin E V, Boyko V P, Maiss E, Frotschl R, Lunina N A, Atabekov J G. Beet yellows closterovirus: complete genome structure and identification of a leader papain-like thiol protease. Virology. 1994;198:311–324. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
