

Tissue culture-specific expression of a naturally occurring tobacco feedback-insensitive anthranilate synthase
- PMID: 9625706
- PMCID: PMC34973
- DOI: 10.1104/pp.117.2.533
Tissue culture-specific expression of a naturally occurring tobacco feedback-insensitive anthranilate synthase
Abstract
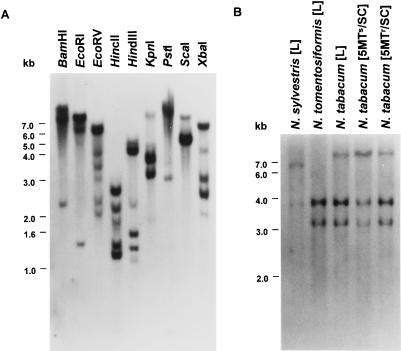
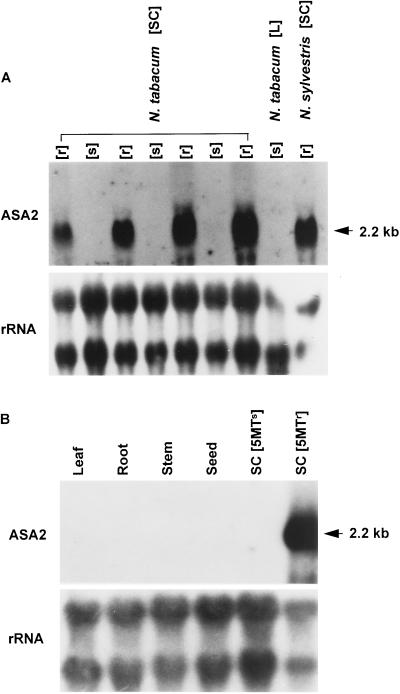
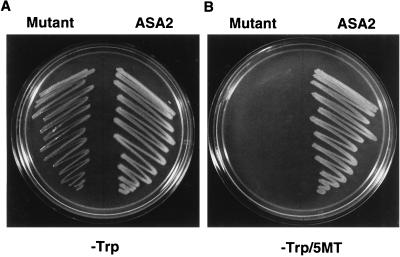
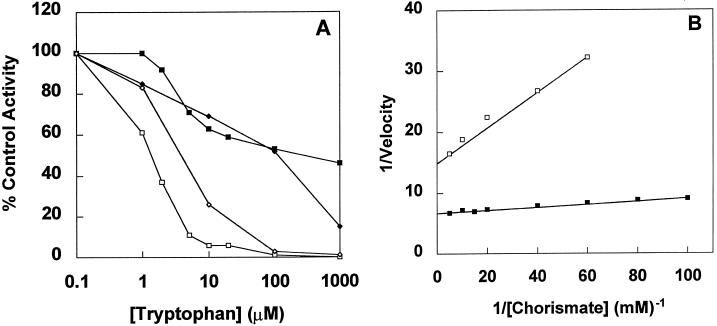
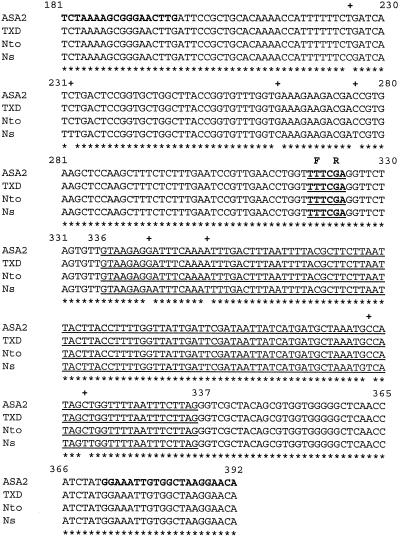
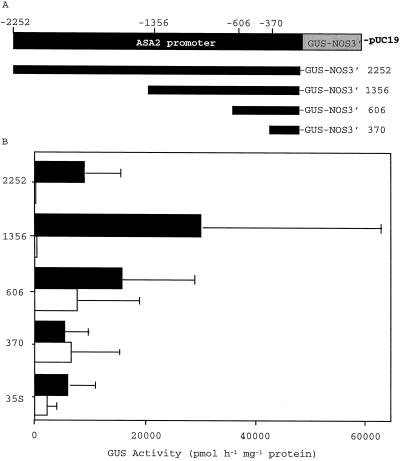
A cDNA and corresponding promoter region for a naturally occurring, feedback-insensitive anthranilate synthase (AS) alpha-subunit gene, ASA2, has been isolated from an unselected, but 5-methyl-tryptophan-resistant (5MTr), tobacco (Nicotiana tabacum) cell line (AB15-12-1). The ASA2 cDNA contains a putative transit peptide sequence, and Southern hybridization shows that more than one closely related sequence is present in the tobacco genome. The ASA2 cDNA complemented a trpE nonsense mutant Escherichia coli strain, allowing growth on 300 microm 5MT-containing minimal medium without tryptophan, and cell extracts contained feedback-insensitive AS activity. The 5MTr was lost when the E. coli strain was transformed with an ASA2 site-directed mutant (phenylalanine-107-arginine-108 --> serine-107-glutamine-108). Identical nucleotide sequences encoding the phenylalanine-107-arginine-108 region have been found in polymerase chain reaction-amplified 326-bp ASA2 genomic fragments of wild-type (5-methyl-tryptophan-sensitive [5MTs]) tobacco and a progenitor species. High-level ASA2 transcriptional expression was detected only in 5MTr-cultured cells, not in 5MTs cells or in plants. Promoter studies indicate that tissue specificity of ASA2 is controlled by the promoter region between -2252 and -607. Since the ASA2 promoter sequences are not substantially different in the 5MTr and 5MTs lines, the increased levels of ASA2 mRNA in the 5MTr lines are most likely due to changes in a regulatory gene affecting ASA2 expression.
Figures



Similar articles
-
Increasing tryptophan synthesis in a forage legume Astragalus sinicus by expressing the tobacco feedback-insensitive anthranilate synthase (ASA2) gene.Plant Physiol. 2000 Jul;123(3):1069-76. doi: 10.1104/pp.123.3.1069. Plant Physiol. 2000. PMID: 10889256 Free PMC article.
-
Use of the tobacco feedback-insensitive anthranilate synthase gene (ASA2) as a selectable marker for legume hairy root transformation.Plant Cell Rep. 2004 Aug;23(1-2):104-13. doi: 10.1007/s00299-004-0789-8. Epub 2004 May 28. Plant Cell Rep. 2004. PMID: 15168071
-
Targeting a nuclear anthranilate synthase alpha-subunit gene to the tobacco plastid genome results in enhanced tryptophan biosynthesis. Return of a gene to its pre-endosymbiotic origin.Plant Physiol. 2001 Sep;127(1):131-41. doi: 10.1104/pp.127.1.131. Plant Physiol. 2001. PMID: 11553741 Free PMC article.
-
Use of 4-methylindole or 7-methyl-DL-tryptophan in a transformant selection system based on the feedback-insensitive anthranilate synthase alpha-subunit of tobacco (ASA2).Plant Cell Rep. 2008 Mar;27(3):509-17. doi: 10.1007/s00299-007-0480-y. Epub 2007 Nov 30. Plant Cell Rep. 2008. PMID: 18060408
-
Overexpression of the feedback-insensitive anthranilate synthase gene in tobacco causes tryptophan accumulation.Plant Cell Rep. 2005 Jan;23(8):548-56. doi: 10.1007/s00299-004-0849-0. Epub 2004 Sep 16. Plant Cell Rep. 2005. PMID: 15375628
Cited by
-
Evolution of aromatic amino acid metabolism in plants: a key driving force behind plant chemical diversity in aromatic natural products.Philos Trans R Soc Lond B Biol Sci. 2024 Nov 18;379(1914):20230352. doi: 10.1098/rstb.2023.0352. Epub 2024 Sep 30. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 39343022 Free PMC article. Review.
-
Characterization of tryptophan-overproducing potato transgenic for a mutant rice anthranilate synthase alpha-subunit gene (OASA1D).Planta. 2005 Oct;222(3):535-45. doi: 10.1007/s00425-005-1565-x. Epub 2005 May 24. Planta. 2005. PMID: 15912354
-
Deregulation of phenylalanine biosynthesis evolved with the emergence of vascular plants.Plant Physiol. 2022 Jan 20;188(1):134-150. doi: 10.1093/plphys/kiab454. Plant Physiol. 2022. PMID: 34633048 Free PMC article.
-
Expression of a feedback insensitive anthranilate synthase gene from tobacco increases free tryptophan in soybean plants.Plant Cell Rep. 2007 Oct;26(10):1763-71. doi: 10.1007/s00299-007-0381-0. Epub 2007 Jun 14. Plant Cell Rep. 2007. PMID: 17569051
-
Emerging trends in research on spatial and temporal organization of terpenoid indole alkaloid pathway in Catharanthus roseus: a literature update.Protoplasma. 2012 Apr;249(2):255-68. doi: 10.1007/s00709-011-0291-4. Epub 2011 Jun 1. Protoplasma. 2012. PMID: 21630129 Review.
References
-
- Altschul F, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. Green and. New York: Wiley-Interscience; 1989.
-
- Bohlmann J, De Luca V, Eilert U, Martin W. Purification and cDNA cloning of anthranilate synthase from Ruta graveolens: modes of expression and properties of native and recombinant enzymes. Plant J. 1995;7:491–501. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
