

The chloroplast atpA gene cluster in Chlamydomonas reinhardtii. Functional analysis of a polycistronic transcription unit
- PMID: 9625716
- PMCID: PMC34983
- DOI: 10.1104/pp.117.2.629
The chloroplast atpA gene cluster in Chlamydomonas reinhardtii. Functional analysis of a polycistronic transcription unit
Abstract
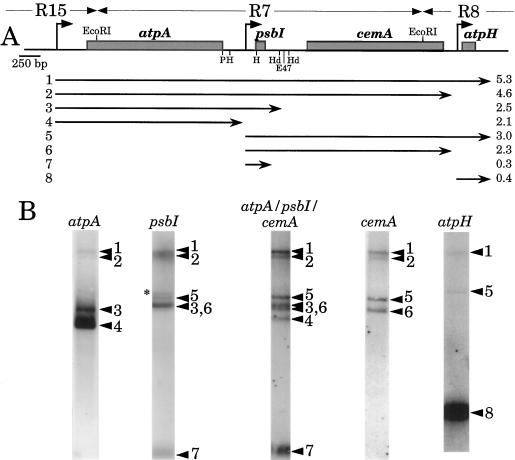
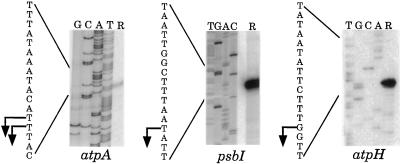
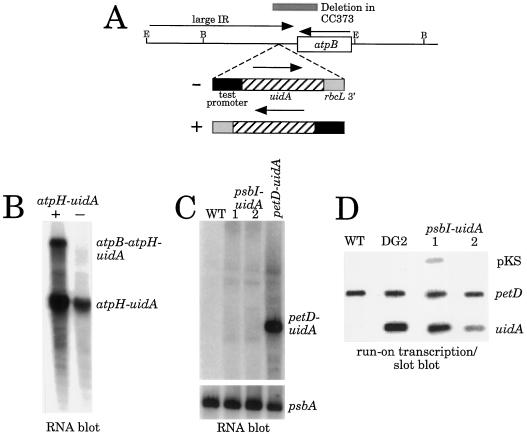
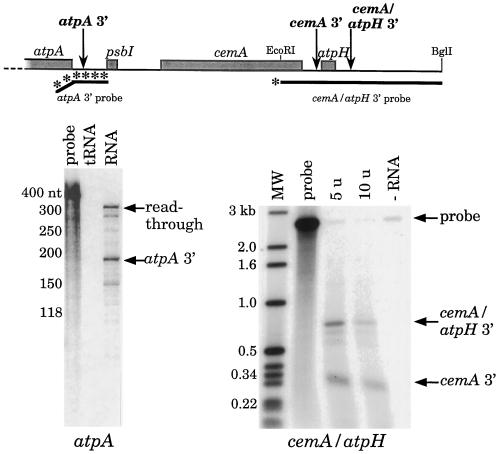
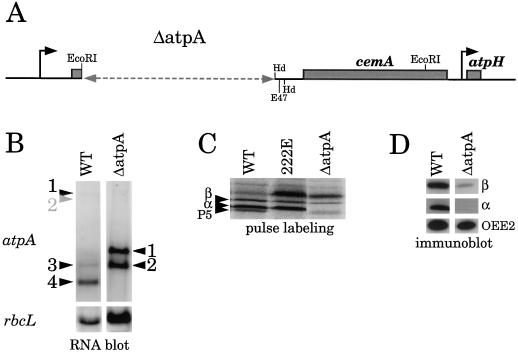
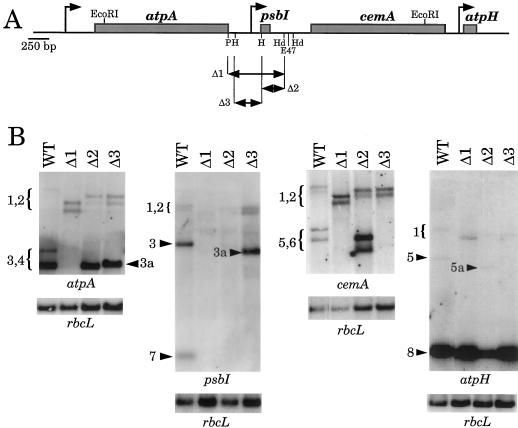
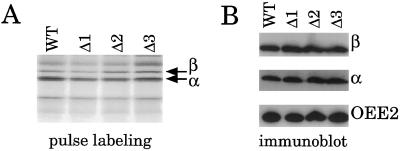
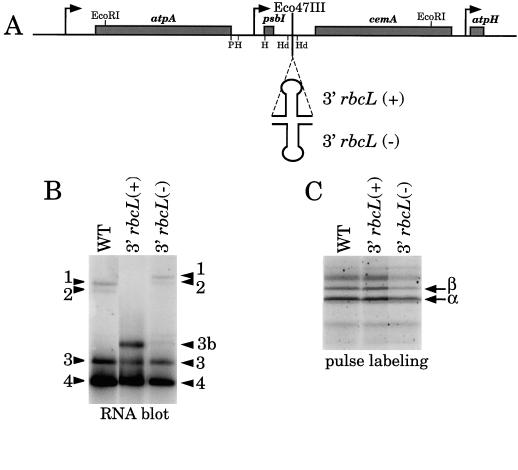
Most chloroplast genes in vascular plants are organized into polycistronic transcription units, which generate a complex pattern of mono-, di-, and polycistronic transcripts. In contrast, most Chlamydomonas reinhardtii chloroplast transcripts characterized to date have been monocistronic. This paper describes the atpA gene cluster in the C. reinhardtii chloroplast genome, which includes the atpA, psbI, cemA, and atpH genes, encoding the alpha-subunit of the coupling-factor-1 (CF1) ATP synthase, a small photosystem II polypeptide, a chloroplast envelope membrane protein, and subunit III of the CF0 ATP synthase, respectively. We show that promoters precede the atpA, psbI, and atpH genes, but not the cemA gene, and that cemA mRNA is present only as part of di-, tri-, or tetracistronic transcripts. Deletions introduced into the gene cluster reveal, first, that CF1-alpha can be translated from di- or polycistronic transcripts, and, second, that substantial reductions in mRNA quantity have minimal effects on protein synthesis rates. We suggest that posttranscriptional mRNA processing is common in C. reinhardtii chloroplasts, permitting the expression of multiple genes from a single promoter.
Figures
Similar articles
-
Target and specificity of a nuclear gene product that participates in mRNA 3'-end formation in Chlamydomonas chloroplasts.J Biol Chem. 1999 Dec 10;274(50):35955-62. doi: 10.1074/jbc.274.50.35955. J Biol Chem. 1999. PMID: 10585484
-
Dual functions of the nucleus-encoded factor TDA1 in trapping and translation activation of atpA transcripts in Chlamydomonas reinhardtii chloroplasts.Plant J. 2011 Sep;67(6):1055-66. doi: 10.1111/j.1365-313X.2011.04657.x. Epub 2011 Jul 18. Plant J. 2011. PMID: 21623973
-
Cotranscription of the wild-type chloroplast atpE gene encoding the CF1/CF0 epsilon subunit with the 3' half of the rps7 gene in Chlamydomonas reinhardtii and characterization of frameshift mutations in atpE.Mol Gen Genet. 1990 Apr;221(2):155-63. doi: 10.1007/BF00261715. Mol Gen Genet. 1990. PMID: 2196429
-
The Chloroplast of Chlamydomonas reinhardtii as a Testbed for Engineering Nitrogen Fixation into Plants.Int J Mol Sci. 2021 Aug 16;22(16):8806. doi: 10.3390/ijms22168806. Int J Mol Sci. 2021. PMID: 34445505 Free PMC article. Review.
-
The Chloroplast Trans-Splicing RNA-Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii.Cells. 2021 Feb 1;10(2):290. doi: 10.3390/cells10020290. Cells. 2021. PMID: 33535503 Free PMC article. Review.
Cited by
-
Integration of chloroplast nucleic acid metabolism into the phosphate deprivation response in Chlamydomonas reinhardtii.Plant Cell. 2007 Mar;19(3):1023-38. doi: 10.1105/tpc.106.045427. Epub 2007 Mar 9. Plant Cell. 2007. PMID: 17351118 Free PMC article.
-
TCA1, a single nuclear-encoded translational activator specific for petA mRNA in Chlamydomonas reinhardtii chloroplast.Genetics. 2001 Sep;159(1):119-32. doi: 10.1093/genetics/159.1.119. Genetics. 2001. PMID: 11560891 Free PMC article.
-
Molecular identification and function of cis- and trans-acting determinants for petA transcript stability in Chlamydomonas reinhardtii chloroplasts.Mol Cell Biol. 2008 Sep;28(17):5529-42. doi: 10.1128/MCB.02056-07. Epub 2008 Jun 23. Mol Cell Biol. 2008. PMID: 18573878 Free PMC article.
-
Chlamydomonas reinhardtii at the crossroads of genomics.Eukaryot Cell. 2003 Dec;2(6):1137-50. doi: 10.1128/EC.2.6.1137-1150.2003. Eukaryot Cell. 2003. PMID: 14665449 Free PMC article. Review. No abstract available.
-
High-throughput sequencing of the chloroplast and mitochondrion of Chlamydomonas reinhardtii to generate improved de novo assemblies, analyze expression patterns and transcript speciation, and evaluate diversity among laboratory strains and wild isolates.Plant J. 2018 Feb;93(3):545-565. doi: 10.1111/tpj.13788. Epub 2018 Jan 7. Plant J. 2018. PMID: 29172250 Free PMC article.
References
-
- Ausubel FM, Brent R, Kingston RG, Moore DD, Seidman JG, Smith JA, Struhl K (1990) Current Protocols in Molecular Biology. Green Publishing Associates and Wiley Interscience, New York
-
- Blowers AD, Klein U, Ellmore GS, Bogorad L. Functional in vivo analyses of the 3′ flanking sequences of the Chlamydomonas chloroplast rbcL and psaB genes. Mol Gen Genet. 1993;238:339–349. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
