

Mechanisms underlying reduced maximum shortening velocity during fatigue of intact, single fibres of mouse muscle
- PMID: 9625883
- PMCID: PMC2231032
- DOI: 10.1111/j.1469-7793.1998.269bz.x
Mechanisms underlying reduced maximum shortening velocity during fatigue of intact, single fibres of mouse muscle
Abstract
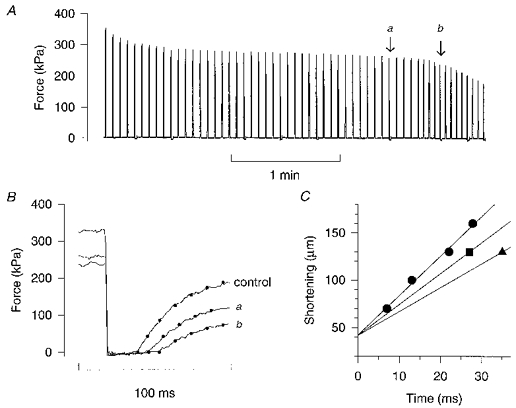
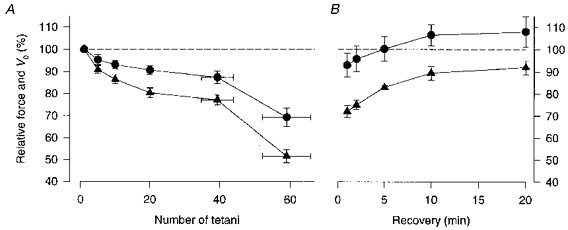
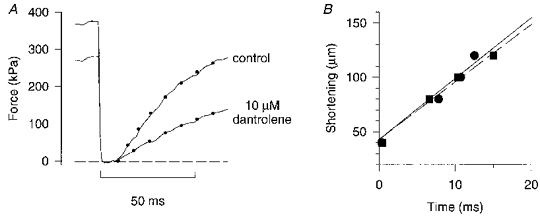
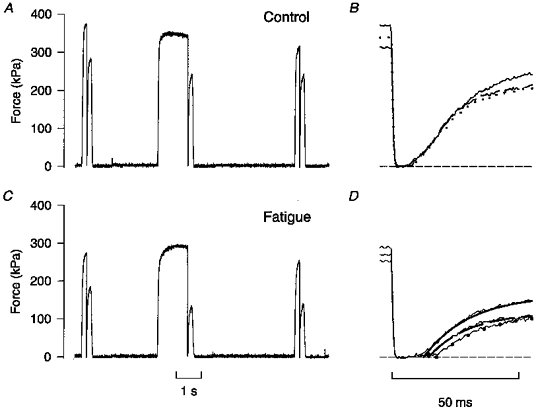
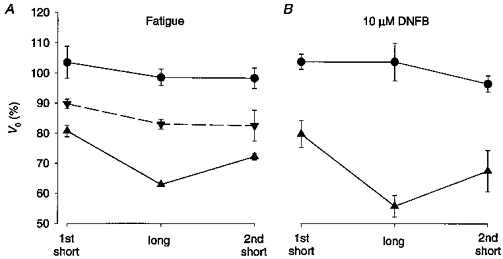
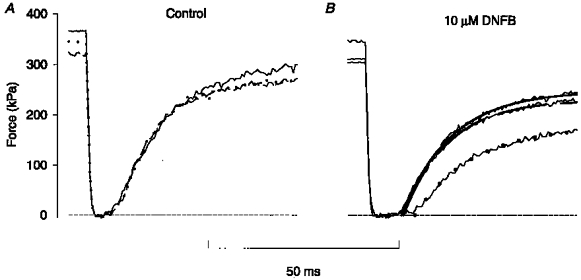
1. The mechanism behind the reduction in shortening velocity in skeletal muscle fatigue is unclear. In the present study we have measured the maximum shortening velocity (V0) with slack tests during fatigue produced by repeated, 350 ms tetani in intact, single muscle fibres from the mouse. We have focused on two possible mechanisms behind the reduction in V0: reduced tetanic Ca2+ and accumulation of ADP. 2. During fatigue V0 initially declined slowly, reaching 90 % of the control after about forty tetani. The rate of decline then increased and V0 fell to 70 % of the control in an additional twenty tetani. The reduction in isometric force followed a similar pattern. 3. Exposing unfatigued fibres to 10 microM dantrolene, which reduces tetanic Ca2+, lowered force by about 35 % but had no effect on V0. 4. In order to see if ADP might increase rapidly during ongoing contractions, we used a protocol with a tetanus of longer duration bracketed by standard-duration tetani. V0 in these three tetani were not significantly different in control, whereas V0 was markedly lower in the longer tetanus during fatigue and in unfatigued fibres where the creatine kinase reaction was inhibited by 10 microM dinitrofluorobenzene. 5. We conclude that the reduction in V0 during fatigue is mainly due to a transient accumulation of ADP, which develops during contractions in fibres with impaired phosphocreatine energy buffering.
Figures
References
-
- Allen DG, Lännergren J, Westerblad H. Muscle cell function during prolonged activity: cellular mechanisms of fatigue. Experimental Physiology. 1995;80:497–527. - PubMed
-
- Butler TM, Siegman MJ, Mooers SU, Barsotti RJ. Myosin light chain phosphorylation does not modulate cross-bridge cycling rate in mouse skeletal muscle. Science. 1983;220:1167–1169. - PubMed
-
- Chase PB, Kushmerick MJ. Effect of physiological ADP concentrations on contraction of single skinned fibers from rabbit fast and slow muscles. American Journal of Physiology. 1995;268:C480–489. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous