

Mammalian p55CDC mediates association of the spindle checkpoint protein Mad2 with the cyclosome/anaphase-promoting complex, and is involved in regulating anaphase onset and late mitotic events
- PMID: 9628895
- PMCID: PMC2132789
- DOI: 10.1083/jcb.141.6.1393
Mammalian p55CDC mediates association of the spindle checkpoint protein Mad2 with the cyclosome/anaphase-promoting complex, and is involved in regulating anaphase onset and late mitotic events
Abstract
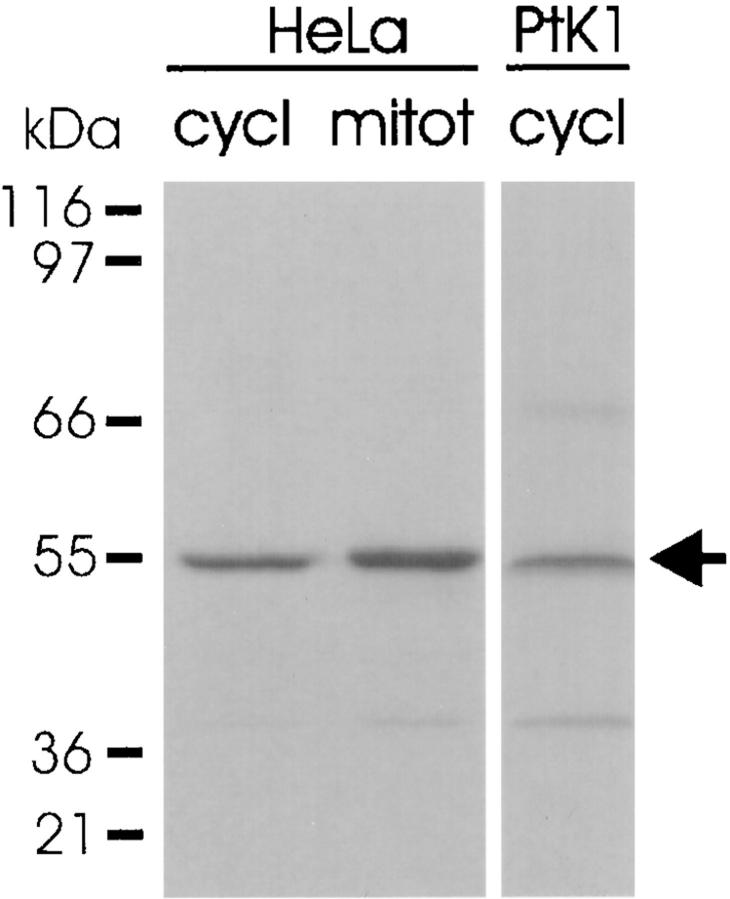
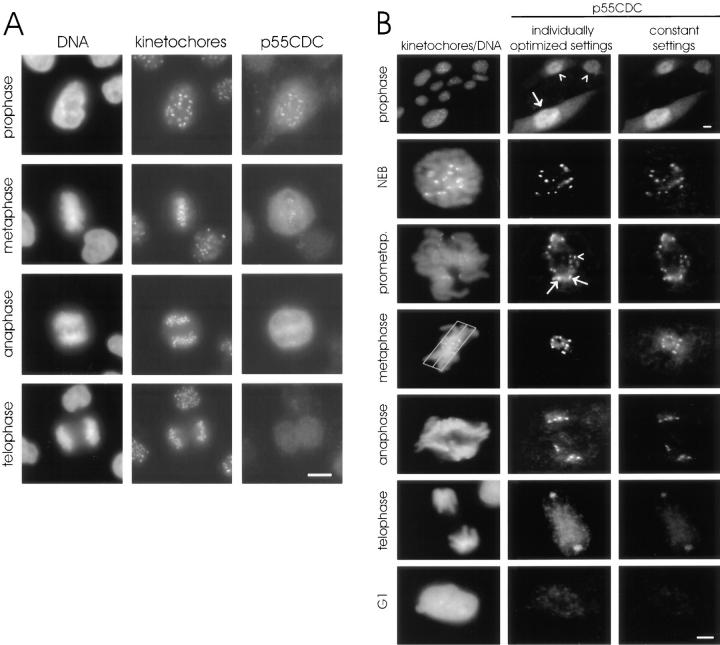
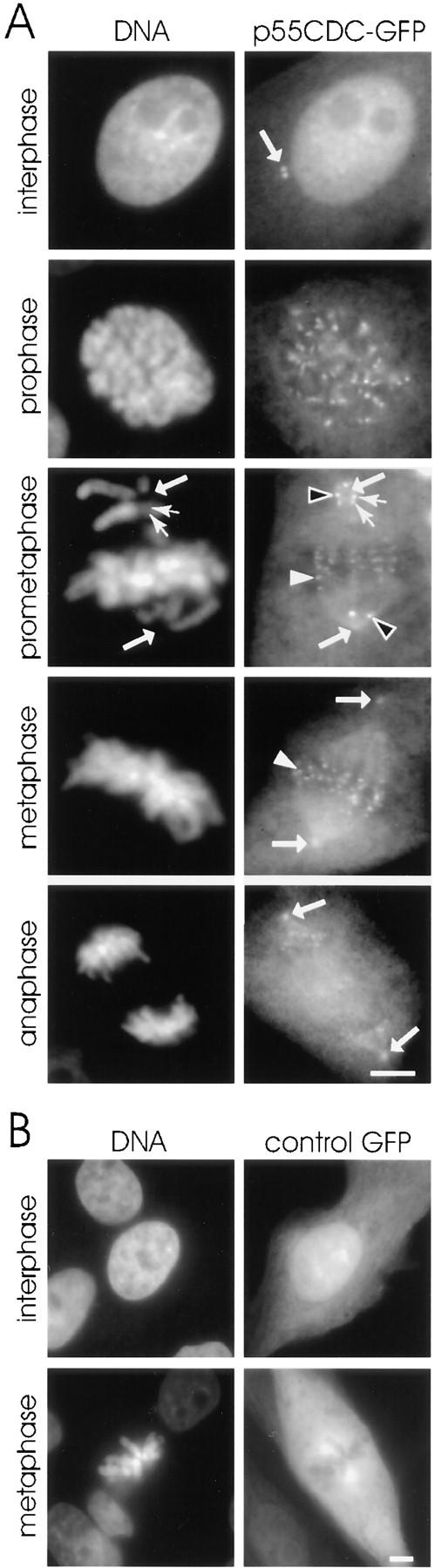
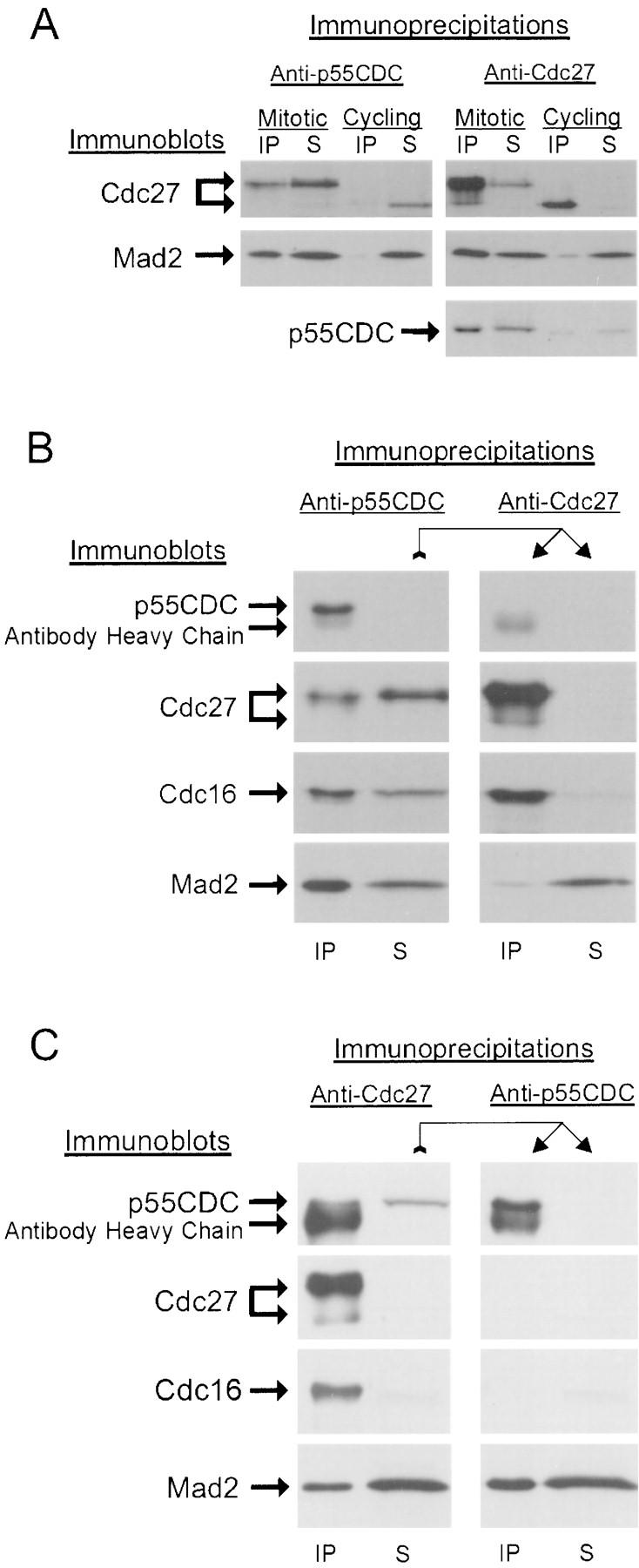
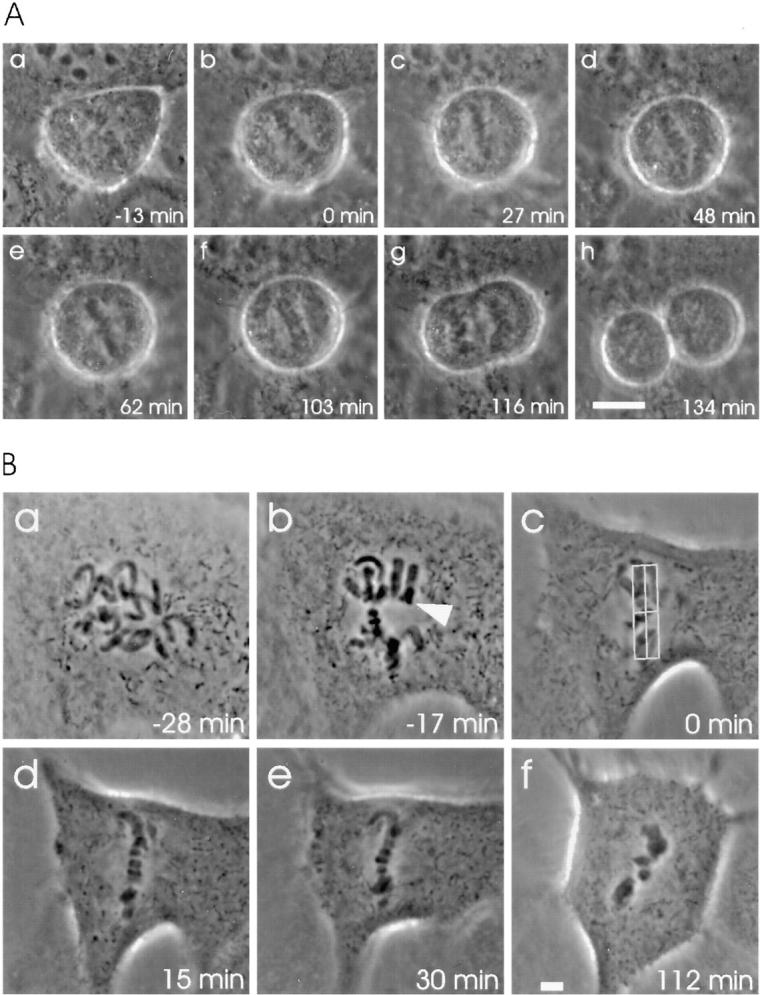
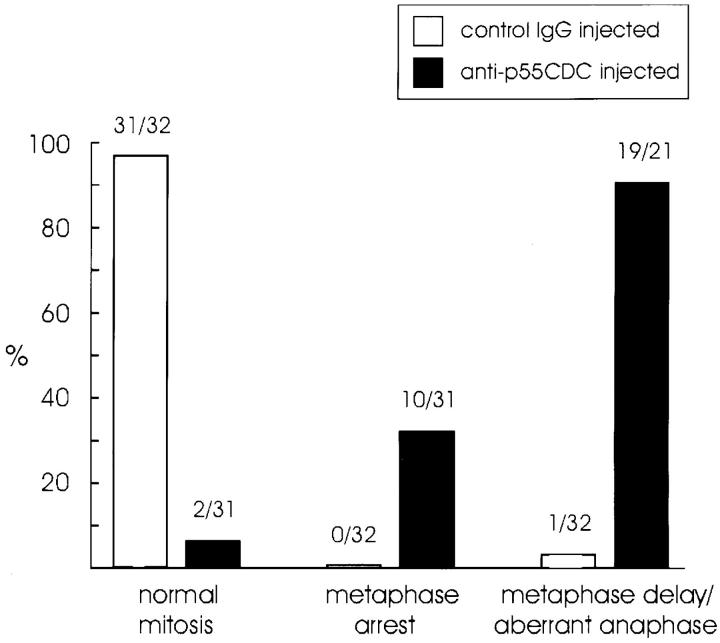
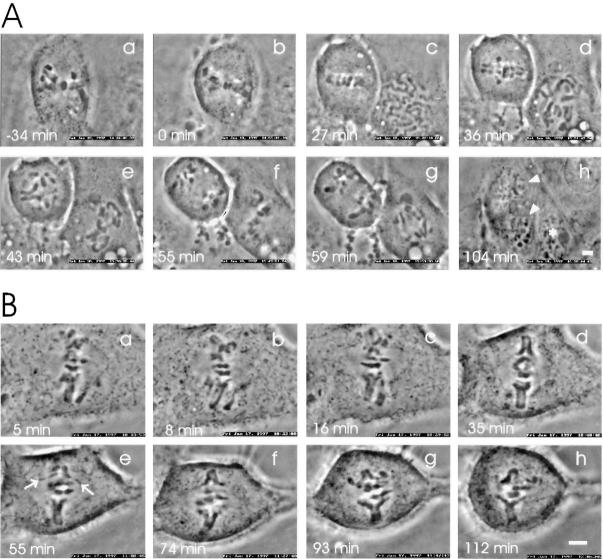
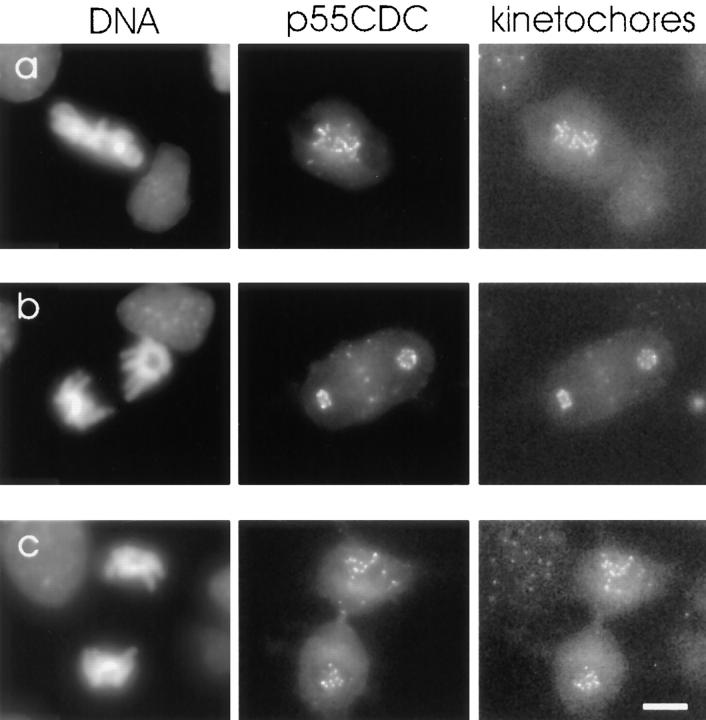
We have investigated the function of p55CDC, a mammalian protein related to Cdc20 and Hct1/Cdh1 in Saccharomyces cerevisiae, and Fizzy and Fizzy-related in Drosophila. Immunofluorescence studies and expression of a p55CDC-GFP chimera demonstrate that p55CDC is concentrated at the kinetochores in M phase cells from late prophase to telophase. Some p55CDC is also associated with the spindle microtubules and spindle poles, and some is diffuse in the cytoplasm. At anaphase, the concentration of p55CDC at the kinetochores gradually diminishes, and is gone by late telophase. In extracts prepared from M phase, but not from interphase HeLa cells, p55CDC coimmunoprecipitates with three important elements of the M phase checkpoint machinery: Cdc27, Cdc16, and Mad2. p55CDC is required for binding Mad2 with the Cdc27 and Cdc16. Thus, it is likely that p55CDC mediates the association of Mad2 with the cyclosome/anaphase-promoting complex. Microinjection of anti-p55CDC antibody into mitotic mammalian cells induces arrest or delay at metaphase, and impairs progression of late mitotic events. These studies suggest that mammalian p55CDC may be part of a regulatory and targeting complex for the anaphase-promoting complex.
Figures
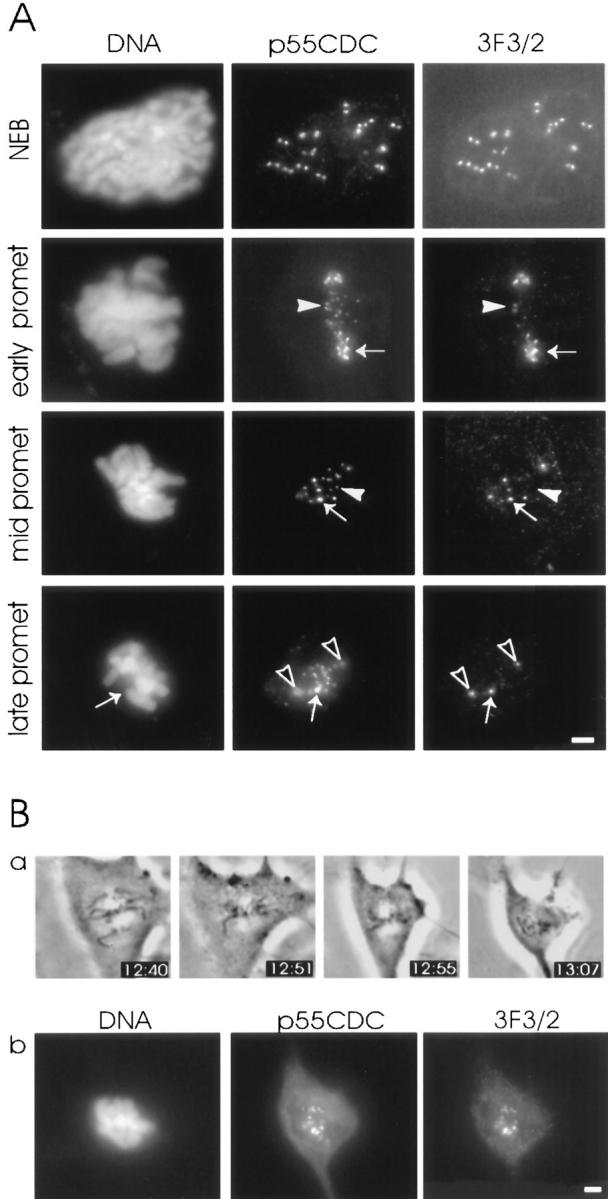
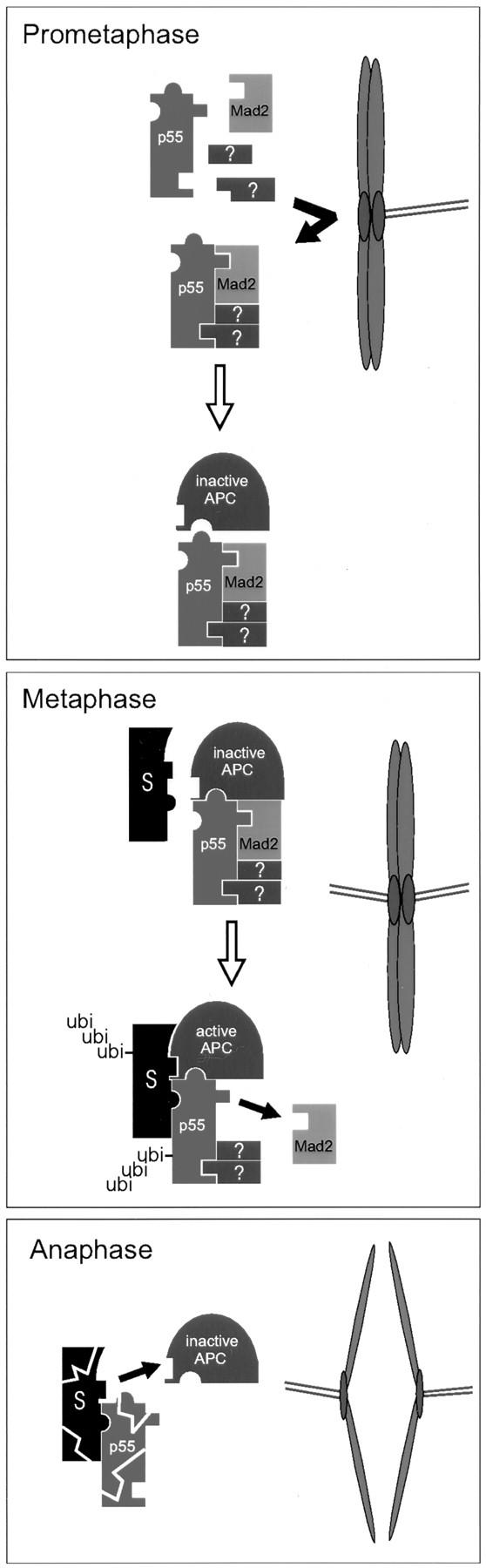
References
-
- Chen R-H, Waters JC, Salmon ED, Murray AW. Association of spindle assembly checkpoint component XMAD2 with unattached kinetochores. Science. 1996;274:242–246. - PubMed
-
- Cohen-Fix O, Peters J-M, Kirschner MW, Koshland D. Anaphase initiation in Saccharomyces cerevisiae is controlled by the APC-dependent degradation of the anaphase inhibitor Pds1p. Genes Dev. 1996;10:3081–3093. - PubMed
-
- Cyert MS, Scherson T, Kirschner MW. Monoclonal antibodies specific for thiophosphorylated proteins recognize XenopusMPF. Dev Biol. 1988;129:209–216. - PubMed
-
- Dawson IA, Roth S, Akam M, Artavanis-Tsakonas S. Mutations of the fizzy locus cause metaphase arrest in Drosophila melanogasterembryos. Development. 1993;117:359–376. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
