

RB and c-Myc activate expression of the E-cadherin gene in epithelial cells through interaction with transcription factor AP-2
- PMID: 9632747
- PMCID: PMC108947
- DOI: 10.1128/MCB.18.7.3647
RB and c-Myc activate expression of the E-cadherin gene in epithelial cells through interaction with transcription factor AP-2
Abstract
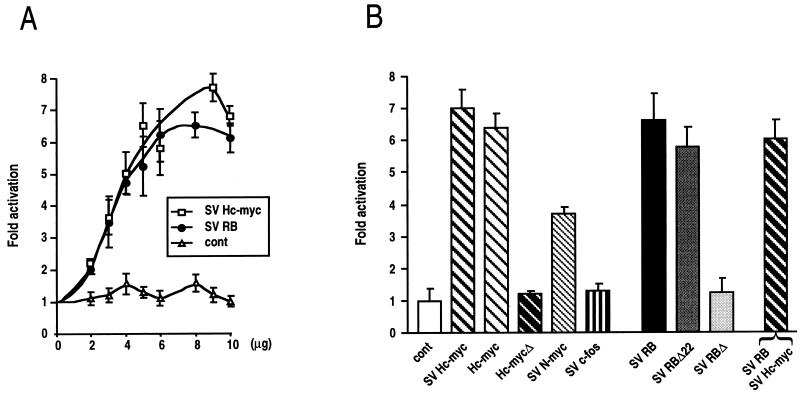
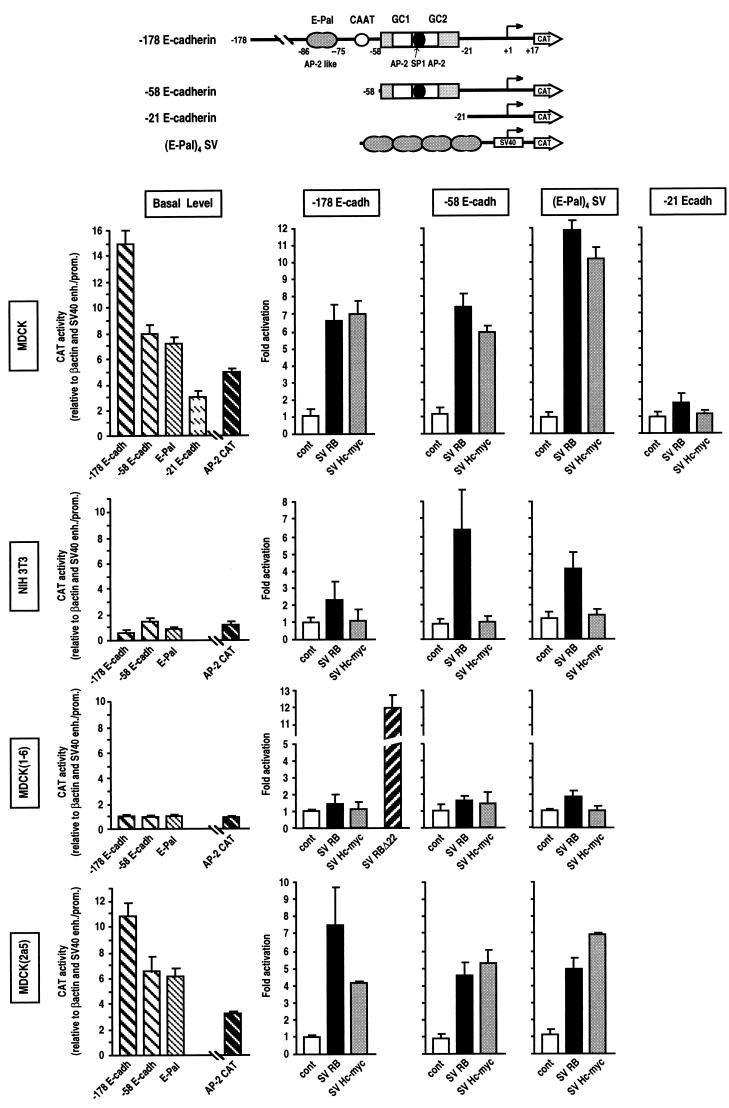
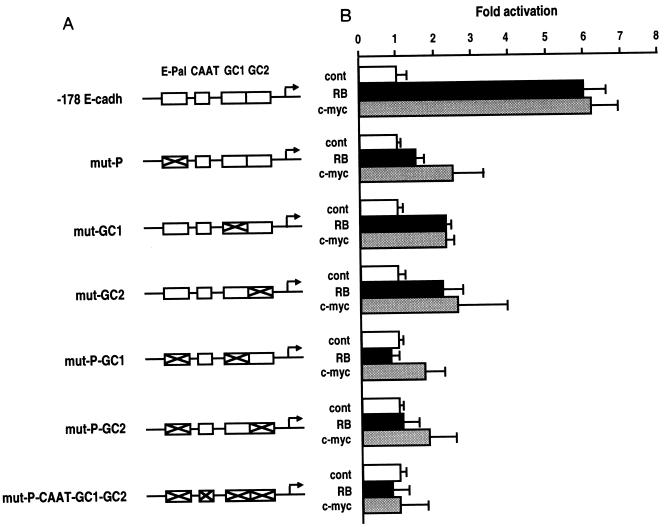
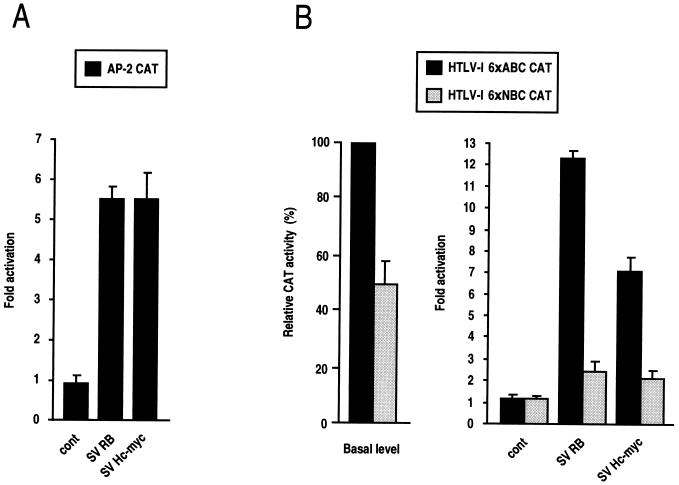
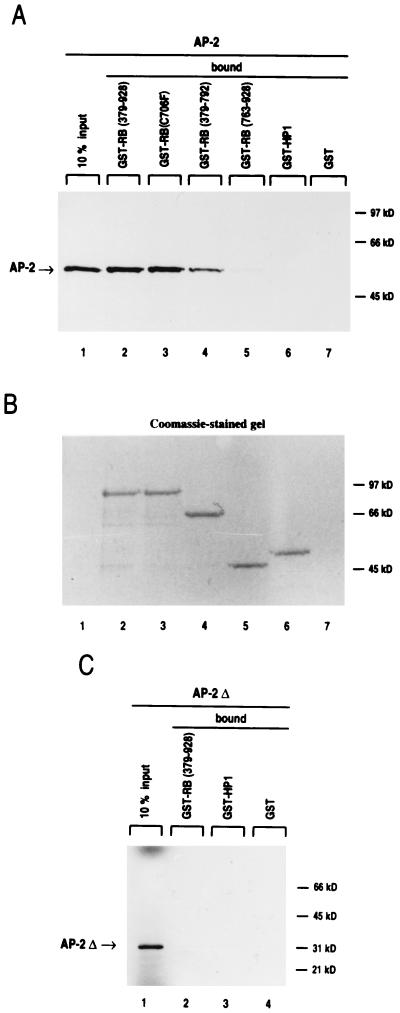
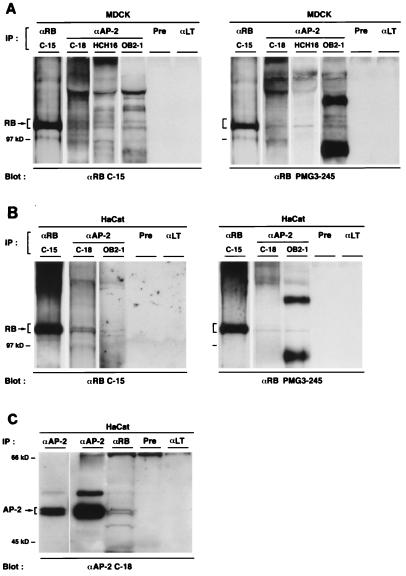
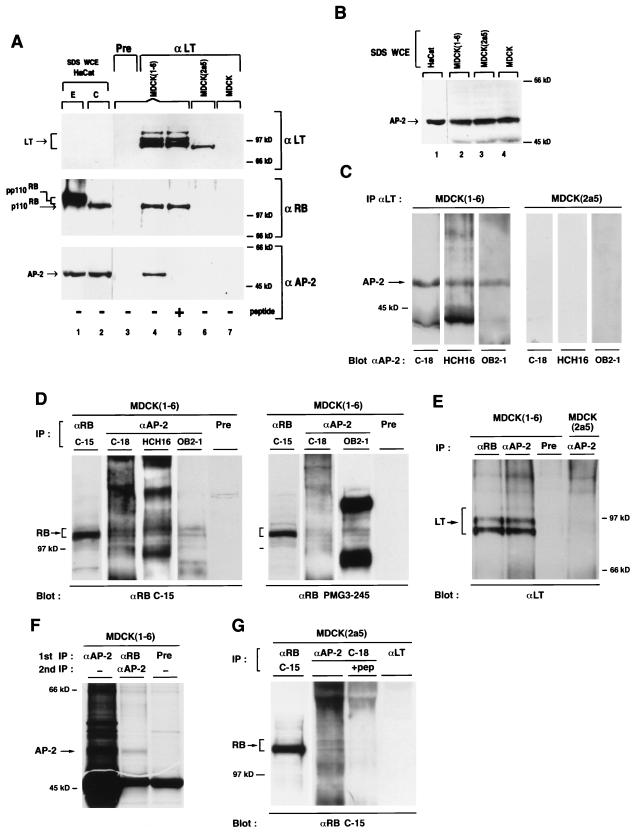
E-cadherin plays a pivotal role in the biogenesis of the first epithelium during development, and its down-regulation is associated with metastasis of carcinomas. We recently reported that inactivation of RB family proteins by simian virus 40 large T antigen (LT) in MDCK epithelial cells results in a mesenchymal conversion associated with invasiveness and a down-regulation of c-Myc. Reexpression of RB or c-Myc in such cells allows the reexpression of epithelial markers including E-cadherin. Here we show that both RB and c-Myc specifically activate transcription of the E-cadherin promoter in epithelial cells but not in NIH 3T3 mesenchymal cells. This transcriptional activity is mediated in both cases by the transcription factor AP-2. In vitro AP-2 and RB interaction involves the N-terminal domain of AP-2 and the oncoprotein binding domain and C-terminal domain of RB. In vivo physical interaction between RB and AP-2 was demonstrated in MDCK and HaCat cells. In LT-transformed MDCK cells, LT, RB, and AP-2 were all coimmunoprecipitated by each of the corresponding antibodies, and a mutation of the RB binding domain of the oncoprotein inhibited its binding to both RB and AP-2. Taken together, our results suggest that there is a tripartite complex between LT, RB, and AP-2 and that the physical and functional interactions between LT and AP-2 are mediated by RB. Moreover, they define RB and c-Myc as coactivators of AP-2 in epithelial cells and shed new light on the significance of the LT-RB complex, linking it to the dedifferentiation processes occurring during tumor progression. These data confirm the important role for RB and c-Myc in the maintenance of the epithelial phenotype and reveal a novel mechanism of gene activation by c-Myc.
Figures
References
-
- Adnane J P, Robbins D. The retinoblastoma susceptibility gene product regulates Myc-mediated transcription. Oncogene. 1995;10:381–387. - PubMed
-
- Batsché E, Lipp M, Crémisi C. Transcriptional repression and activation in the same cell type of the human c-myc promoter by the retinoblastoma gene protein: antagonisation of both effects by SV40 T antigen. Oncogene. 1994;9:2235–2243. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials