

Identification of major binding proteins and substrates for the SH2-containing protein tyrosine phosphatase SHP-1 in macrophages
- PMID: 9632768
- PMCID: PMC108968
- DOI: 10.1128/MCB.18.7.3838
Identification of major binding proteins and substrates for the SH2-containing protein tyrosine phosphatase SHP-1 in macrophages
Abstract
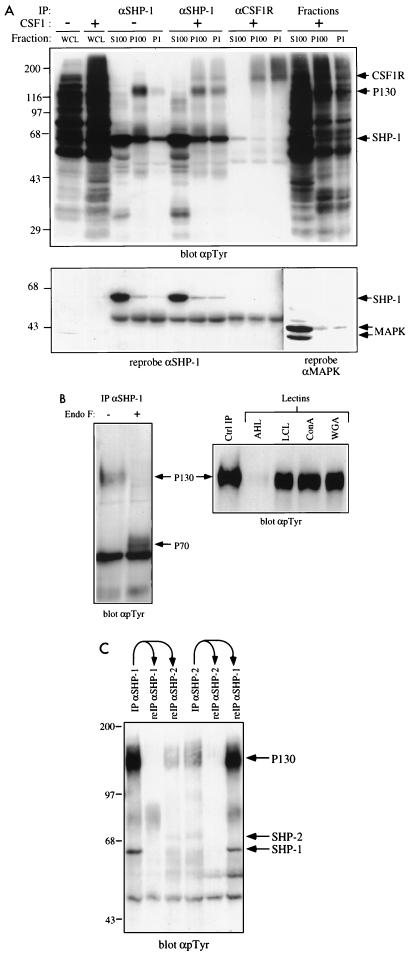
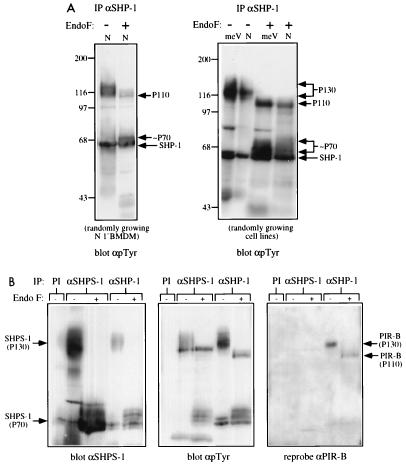
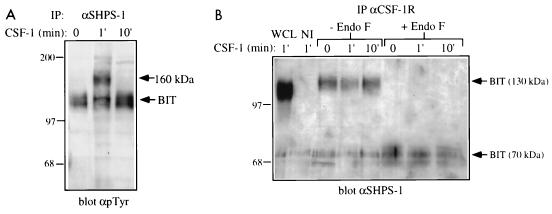
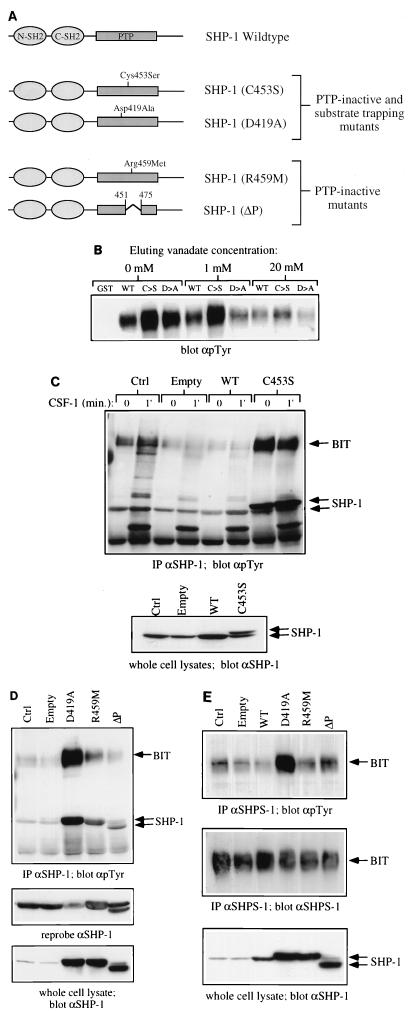
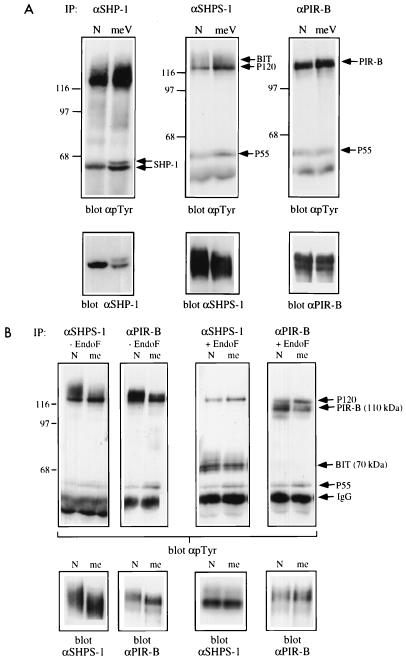
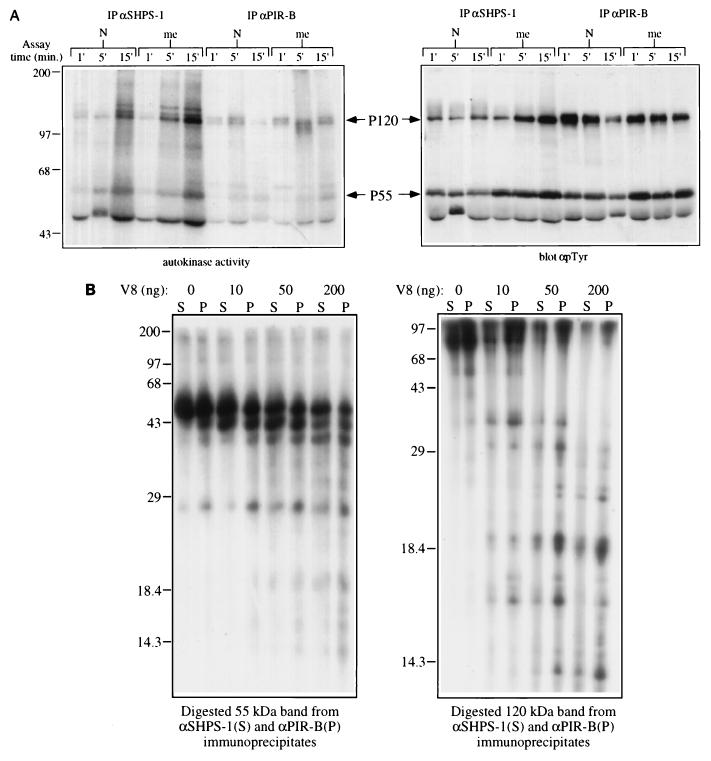
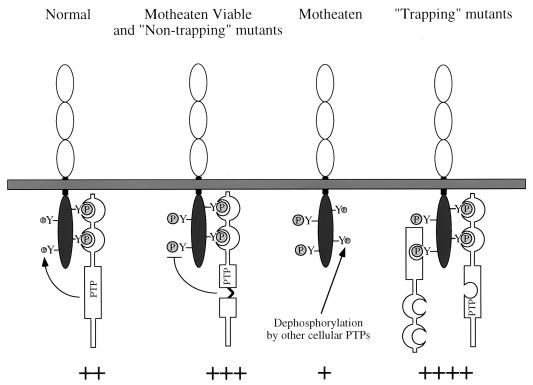
The protein tyrosine phosphatase SHP-1 is a critical regulator of macrophage biology, but its detailed mechanism of action remains largely undefined. SHP-1 associates with a 130-kDa tyrosyl-phosphorylated species (P130) in macrophages, suggesting that P130 might be an SHP-1 regulator and/or substrate. Here we show that P130 consists of two transmembrane glycoproteins, which we identify as PIR-B/p91A and the signal-regulatory protein (SIRP) family member BIT. These proteins also form separate complexes with SHP-2. BIT, but not PIR-B, is in a complex with the colony-stimulating factor 1 receptor (CSF-1R), suggesting that BIT may direct SHP-1 to the CSF-1R. BIT and PIR-B bind preferentially to substrate-trapping mutants of SHP-1 and are hyperphosphorylated in macrophages from motheaten viable mice, which express catalytically impaired forms of SHP-1, indicating that these proteins are SHP-1 substrates. However, BIT and PIR-B are hypophosphorylated in motheaten macrophages, which completely lack SHP-1 expression. These data suggest a model in which SHP-1 dephosphorylates specific sites on BIT and PIR-B while protecting other sites from dephosphorylation via its SH2 domains. Finally, BIT and PIR-B associate with two tyrosyl phosphoproteins and a tyrosine kinase activity. Tyrosyl phosphorylation of these proteins and the level of the associated kinase activity are increased in the absence of SHP-1. Our data suggest that BIT and PIR-B recruit multiple signaling molecules to receptor complexes, where they are regulated by SHP-1 and/or SHP-2.
Figures
Similar articles
-
Regulation of colony-stimulating factor 1 receptor signaling by the SH2 domain-containing tyrosine phosphatase SHPTP1.Mol Cell Biol. 1996 Jul;16(7):3685-97. doi: 10.1128/MCB.16.7.3685. Mol Cell Biol. 1996. PMID: 8668185 Free PMC article.
-
The major SHP-1-binding, tyrosine-phosphorylated protein in macrophages is a member of the KIR/LIR family and an SHP-1 substrate.Oncogene. 1998 Nov 12;17(19):2535-41. doi: 10.1038/sj.onc.1202203. Oncogene. 1998. PMID: 9824165
-
High expression of inhibitory receptor SHPS-1 and its association with protein-tyrosine phosphatase SHP-1 in macrophages.J Biol Chem. 1998 Aug 28;273(35):22719-28. doi: 10.1074/jbc.273.35.22719. J Biol Chem. 1998. PMID: 9712903
-
SHP-1 and SHP-2 in T cells: two phosphatases functioning at many levels.Immunol Rev. 2009 Mar;228(1):342-59. doi: 10.1111/j.1600-065X.2008.00760.x. Immunol Rev. 2009. PMID: 19290938 Free PMC article. Review.
-
Osteoclasts and giant cells: macrophage-macrophage fusion mechanism.Int J Exp Pathol. 2000 Oct;81(5):291-304. doi: 10.1111/j.1365-2613.2000.00164.x. Int J Exp Pathol. 2000. PMID: 11168677 Free PMC article. Review.
Cited by
-
A pan-allelic human SIRPα-blocking antibody, ES004-B5, promotes tumor killing by enhancing macrophage phagocytosis and subsequently inducing an effective T-cell response.Antib Ther. 2024 Aug 28;7(3):266-280. doi: 10.1093/abt/tbae022. eCollection 2024 Jul. Antib Ther. 2024. PMID: 39257438 Free PMC article.
-
Separation and characterization of the activated pool of colony-stimulating factor 1 receptor forming distinct multimeric complexes with signalling molecules in macrophages.Mol Cell Biol. 1999 Jun;19(6):4079-92. doi: 10.1128/MCB.19.6.4079. Mol Cell Biol. 1999. PMID: 10330148 Free PMC article.
-
The SH3-SAM adaptor HACS1 is up-regulated in B cell activation signaling cascades.J Exp Med. 2004 Sep 20;200(6):737-47. doi: 10.1084/jem.20031816. J Exp Med. 2004. PMID: 15381729 Free PMC article.
-
Protein tyrosine phosphatase function: the substrate perspective.Biochem J. 2007 Feb 15;402(1):1-15. doi: 10.1042/BJ20061548. Biochem J. 2007. PMID: 17238862 Free PMC article. Review.
-
On the relationship between extant innate immune receptors and the evolutionary origins of jawed vertebrate adaptive immunity.Immunogenetics. 2022 Feb;74(1):111-128. doi: 10.1007/s00251-021-01232-7. Epub 2022 Jan 4. Immunogenetics. 2022. PMID: 34981186 Review.
References
-
- Adachi M, Fisher E H, Ihle J, Imai K, Jirik F, Neel B, Pawson T, Shen S-H, Thomas M, Ullrich A, Zhao Z. Mammalian SH2-containing protein tyrosine phosphatases. Cell. 1996;85:15. - PubMed
-
- Barford D, Flint A J, Tonks N K. Crystal structure of human protein tyrosine phosphatase 1B. Science. 1994;263:1397–1404. - PubMed
-
- Bignon J S, Siminovitch K A. Identification of PTP1C mutation as a genetic defect in motheaten and viable motheaten mice: a step toward defining the roles of protein tyrosine phosphatases in the regulation of hemopoietic cell differentiation and function. Clin Immunol Immunopathol. 1994;73:168–179. - PubMed
-
- Burshtyn, D. N., and E. O. Long. Regulation through inhibitory receptors: lessons from natural killer cells. Trends Cell Biol., in press. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous