

A novel phosphorylation-dependent RNase activity of GAP-SH3 binding protein: a potential link between signal transduction and RNA stability
- PMID: 9632780
- PMCID: PMC108980
- DOI: 10.1128/MCB.18.7.3956
A novel phosphorylation-dependent RNase activity of GAP-SH3 binding protein: a potential link between signal transduction and RNA stability
Abstract
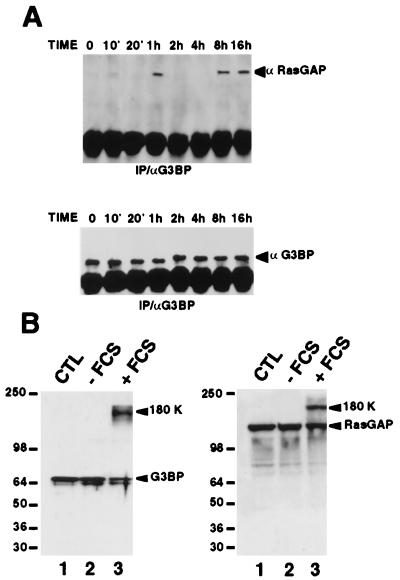
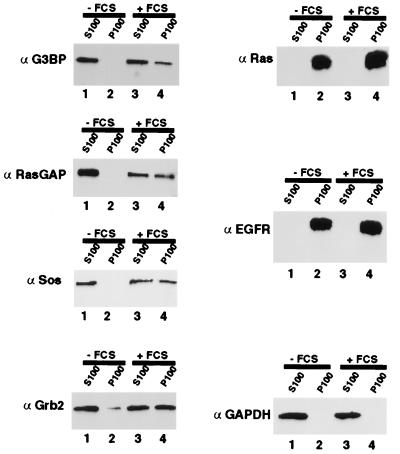
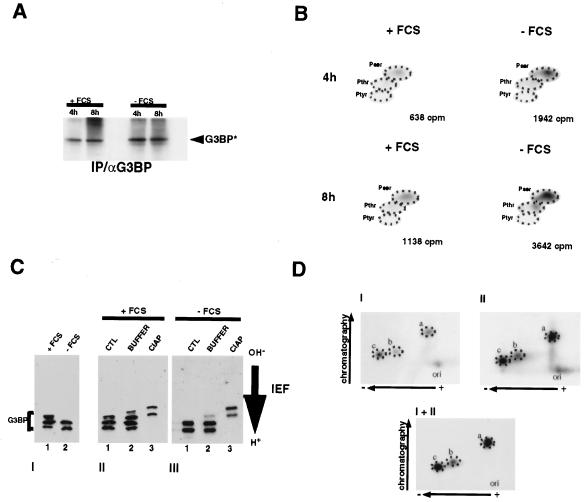
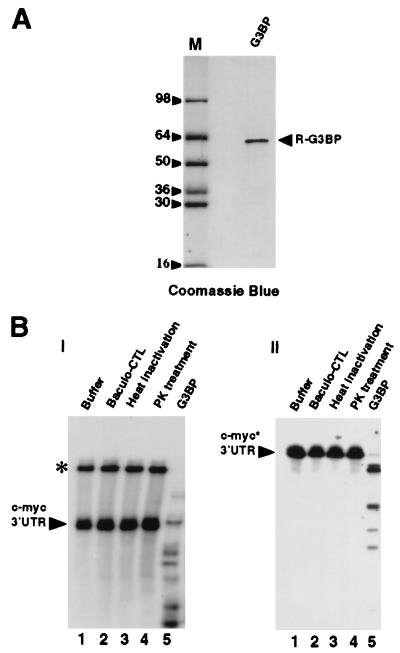
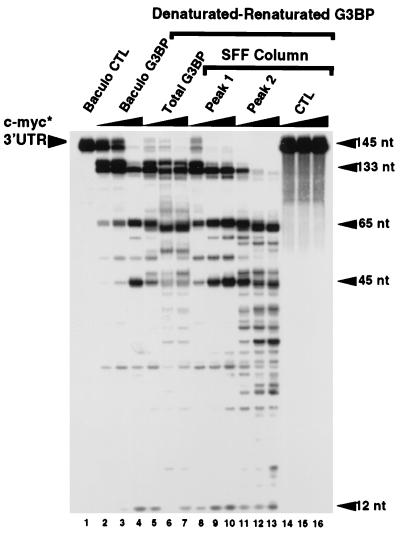
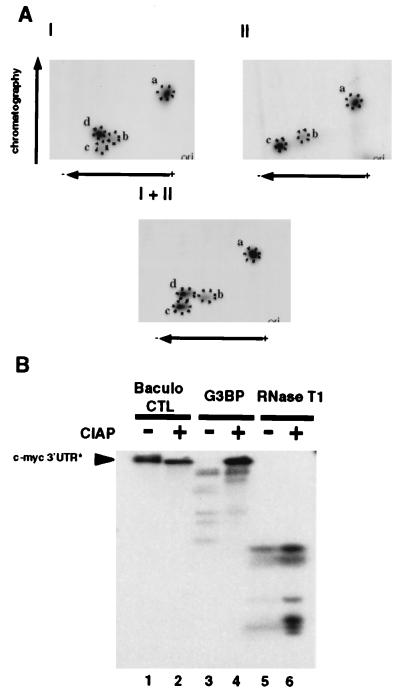
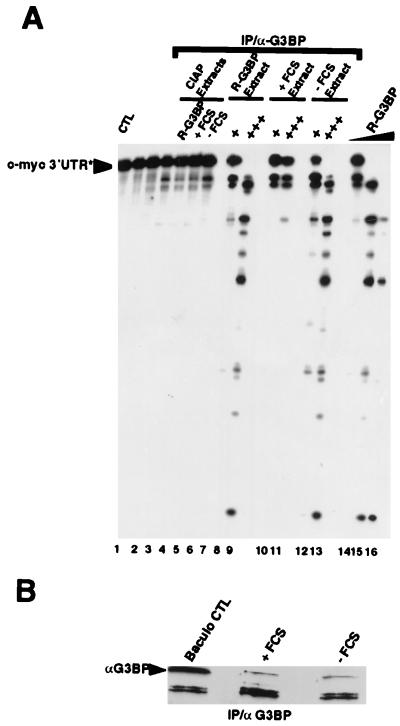
A potential p120 GTPase-activating protein (RasGAP) effector, G3BP (RasGAP Src homology 3 [SH3] binding protein), was previously identified based on its ability to bind the SH3 domain of RasGAP. Here we show that G3BP colocalizes and physically interacts with RasGAP at the plasma membrane of serum-stimulated but not quiescent Chinese hamster lung fibroblasts. In quiescent cells, G3BP was hyperphosphorylated on serine residues, and this modification was essential for its activity. Indeed, G3BP harbors a phosphorylation-dependent RNase activity which specifically cleaves the 3'-untranslated region of human c-myc mRNA. The endoribonuclease activity of G3BP can initiate mRNA degradation and therefore represents a link between a RasGAP-mediated signaling pathway and RNA turnover.
Figures
References
-
- Blanchard J M, Piechaczyk M, Dani C, Chambard J C, Franchl A, Pouyssegur J, Jeanteur P. c-myc gene is transcribed at high rate in G0-arrested fibroblasts and is post-transcriptionally regulated in response to growth factors. Nature. 1985;317:443–445. - PubMed
-
- Boyle W J, van der Geer P, Hunter T. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 1991;201:110–149. - PubMed
-
- Brannan C I, Perkins A S, Vogel K S, Ratner N, Nordlund M L, Reid S W, Buchberg A M, Jenkins N A, Parada L F, Copeland N G. Targeted disruption of the neurofibromatosis type-1 gene leads to developmental abnormalities in heart and various neural crest-derived tissues. Genes Dev. 1994;8:1019–1029. . (Erratum, 8:2792.) - PubMed
-
- Bryant S S, Mitchell A L, Collins F, Miao W, Marshall M, Jove R. N-terminal sequences contained in the Src homology 2 and 3 domains of p120 GTPase-activating protein are required for full catalytic activity toward Ras. J Biol Chem. 1996;271:5195–5199. - PubMed
-
- Buday L, Downward J. Epidermal growth factor regulates p21ras through the formation of a complex of receptor, Grb2 adapter protein, and Sos nucleotide exchange factor. Cell. 1993;73:611–620. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous