

Human cyclin K, a novel RNA polymerase II-associated cyclin possessing both carboxy-terminal domain kinase and Cdk-activating kinase activity
- PMID: 9632813
- PMCID: PMC109013
- DOI: 10.1128/MCB.18.7.4291
Human cyclin K, a novel RNA polymerase II-associated cyclin possessing both carboxy-terminal domain kinase and Cdk-activating kinase activity
Abstract
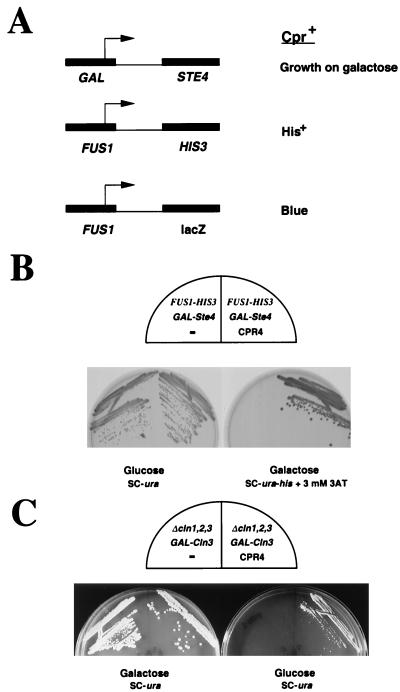
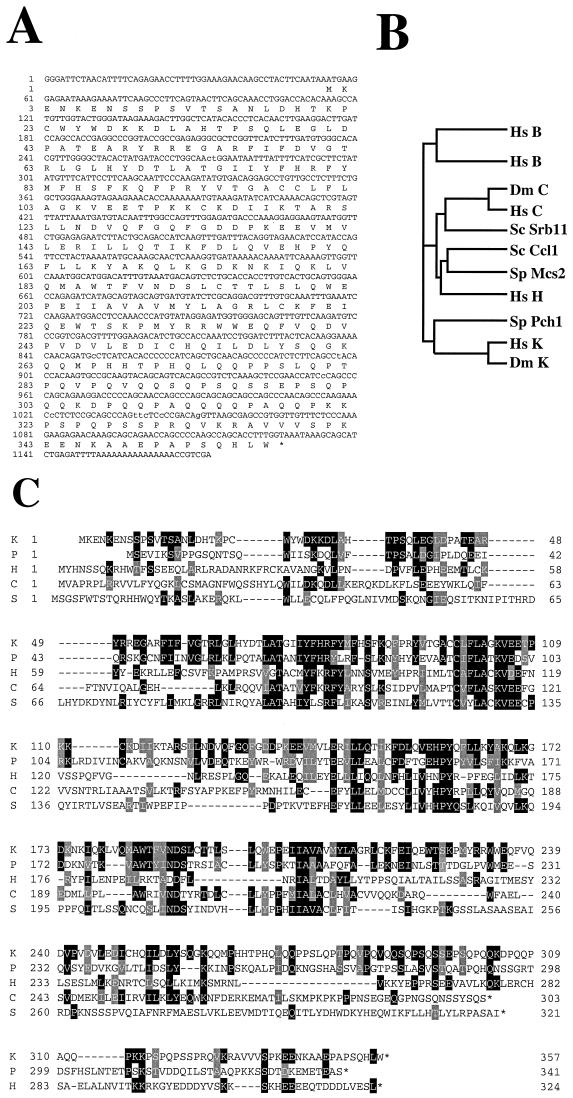
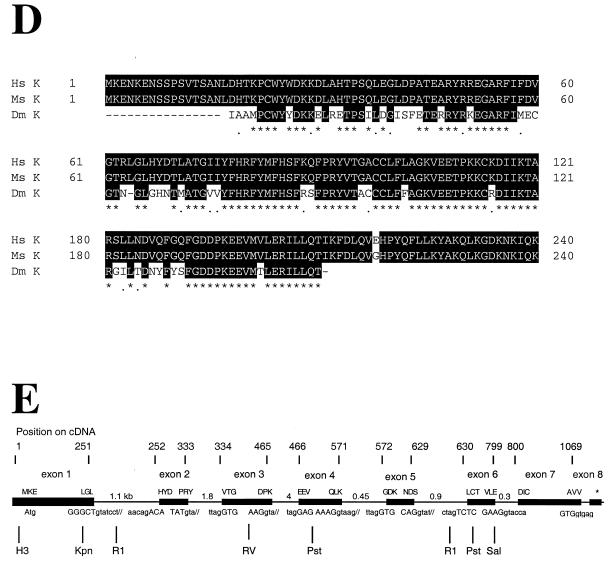
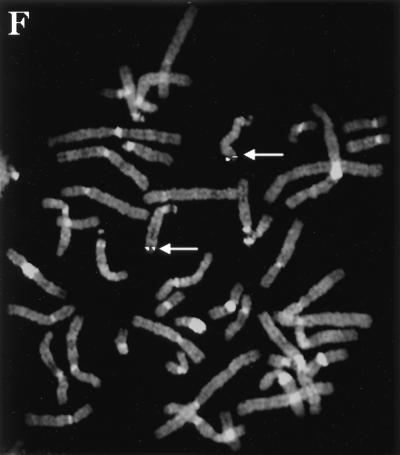
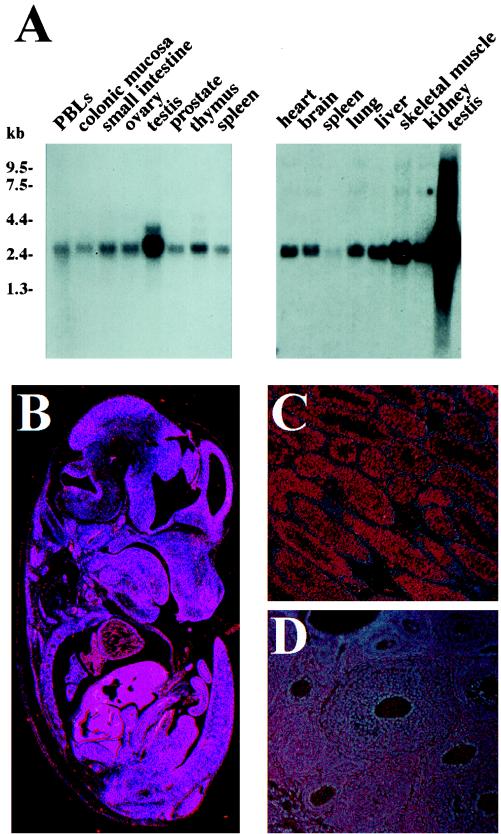
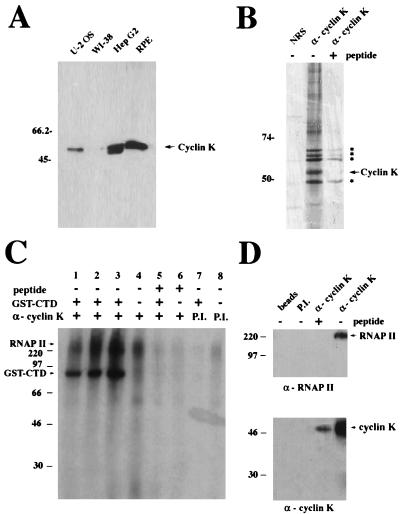
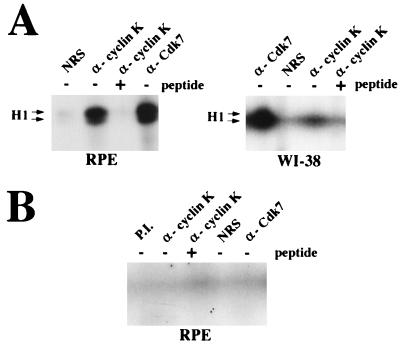
The gene coding for human cyclin K was isolated as a CPR (cell-cycle progression restoration) gene by virtue of its ability to impart a Far- phenotype to the budding yeast Saccharomyces cerevisiae and to rescue the lethality of a deletion of the G1 cyclin genes CLN1, CLN2, and CLN3. The cyclin K gene encodes a 357-amino-acid protein most closely related to human cyclins C and H, which have been proposed to play a role in regulating basal transcription through their association with and activation of cyclin-dependent kinases (Cdks) that phosphorylate the carboxyl-terminal domain (CTD) of the large subunit of RNA polymerase II (RNAP II). Murine and Drosophila melanogaster homologs of cyclin K have also been identified. Cyclin K mRNA is ubiquitously expressed in adult mouse and human tissues, but is most abundant in the developing germ cells of the adult testis and ovaries. Cyclin K is associated with potent CTD kinase and Cdk kinase (CAK) activity in vitro and coimmunoprecipitates with the large subunit of RNAP II. Thus, cyclin K represents a new member of the "transcription" cyclin family which may play a dual role in regulating Cdk and RNAP II activity.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases