

Identification of the amino terminus of neuronal Ca2+ channel alpha1 subunits alpha1B and alpha1E as an essential determinant of G-protein modulation
- PMID: 9634547
- PMCID: PMC6792552
- DOI: 10.1523/JNEUROSCI.18-13-04815.1998
Identification of the amino terminus of neuronal Ca2+ channel alpha1 subunits alpha1B and alpha1E as an essential determinant of G-protein modulation
Abstract
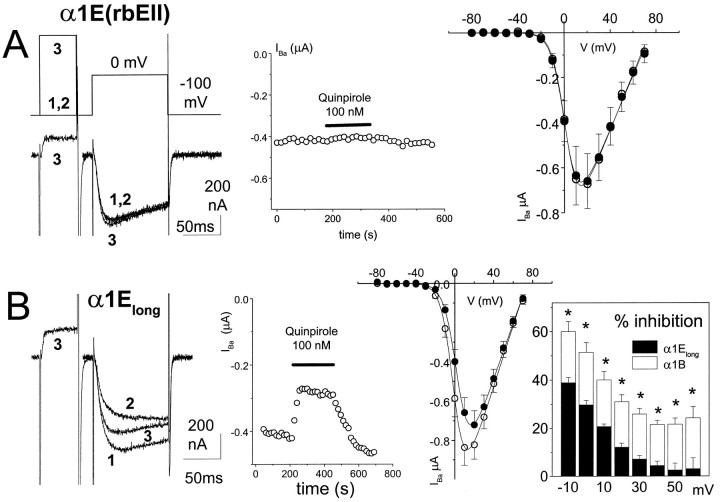
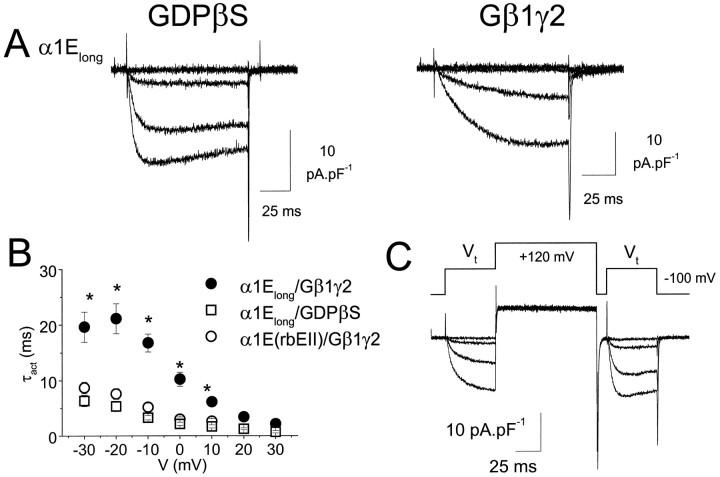
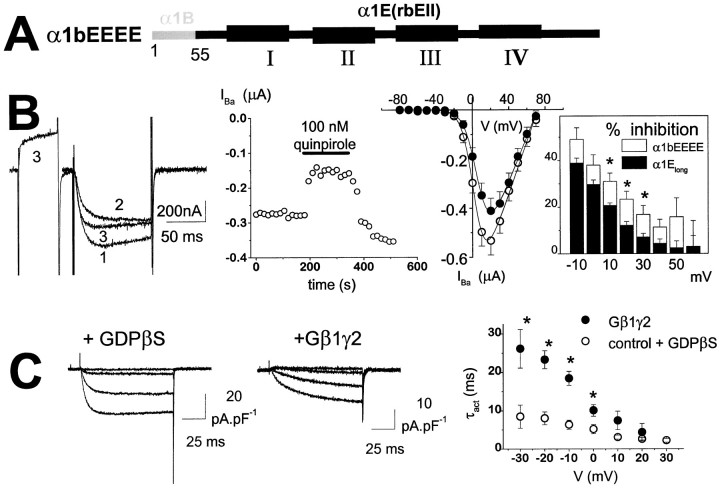
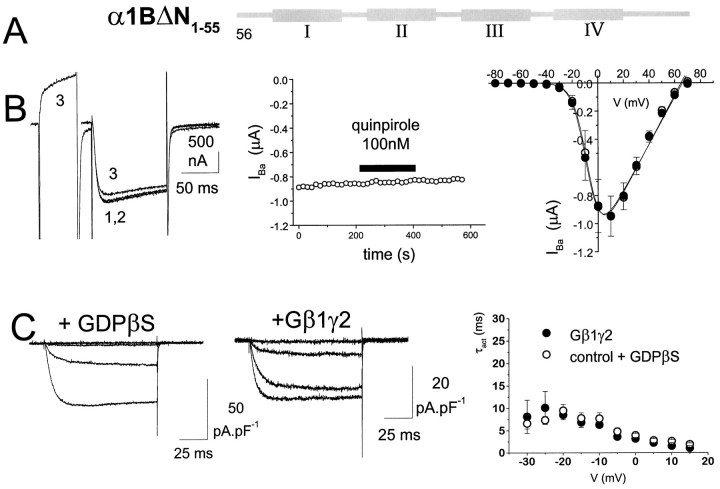
We have examined the basis for G-protein modulation of the neuronal voltage-dependent calcium channels (VDCCs) alpha1E and alpha1B. A novel PCR product of alpha1E was isolated from rat brain. This contained an extended 5' DNA sequence and was subcloned onto the previously cloned isoform rbEII, giving rise to alpha1Elong whose N terminus was extended by 50 amino acids. VDCC alpha1 subunit constructs were co-expressed with the accessory alpha2-delta and beta2a subunits in Xenopus oocytes and mammalian (COS-7) cells. The alpha1Elong showed biophysical properties similar to those of rbEII; however, when G-protein modulation of expressed alpha1 subunits was induced by activation of co-expressed dopamine (D2) receptors with quinpirole (100 nM) in oocytes, or by co-transfection of Gbeta1gamma2 subunits in COS-7 cells, alpha1Elong, unlike alpha1E(rbEII), was found to be G-protein-modulated, in terms of both a slowing of activation kinetics and a reduction in current amplitude. However, alpha1Elong showed less modulation than alpha1B, and substitution of the alpha1E1-50 with the corresponding region of alpha1B1-55 produced a chimera alpha1bEEEE, with G-protein modulation intermediate between alpha1Elong and alpha1B. Furthermore, deletion of the N-terminal 1-55 sequence from alpha1B produced alpha1BDeltaN1-55, which could not be modulated, thus identifying the N-terminal domain as essential for G-protein modulation. Taken together with previous studies, these results indicate that the intracellular N terminus of alpha1E1-50 and alpha1B1-55 is likely to contribute to a multicomponent site, together with the intracellular I-II loop and/or the C-terminal tail, which are involved in Gbetagamma binding and/or in subsequent modulation of channel gating.
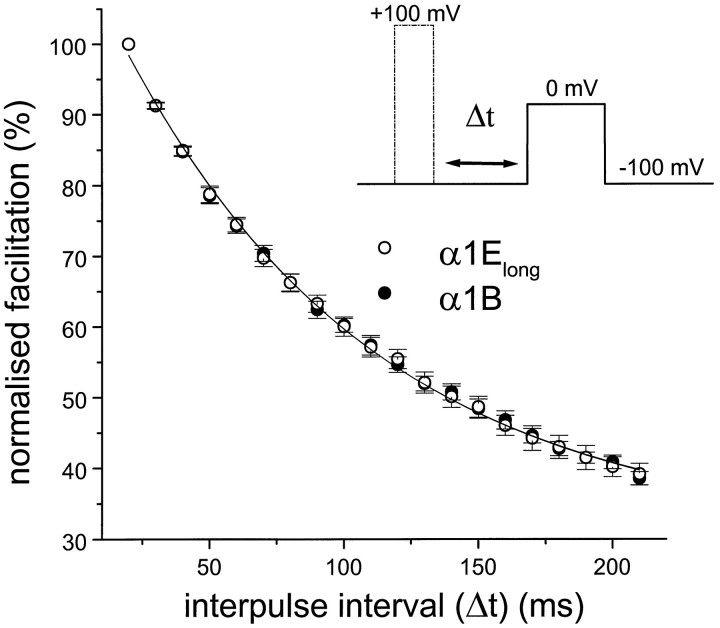
Figures

References
-
- Brice NL, Berrow NS, Campbell V, Page KM, Brickley K, Tedder I, Dolphin AC. Importance of the different β subunits in the membrane expression of the α1A and α2 calcium channel subunits: studies using a depolarisation-sensitive α1A antibody. Eur J Neurosci. 1997;9:749–759. - PubMed
-
- Campbell V, Berrow N, Brickley K, Page K, Wade R, Dolphin AC. Voltage-dependent calcium channel β-subunits in combination with α1 subunits have a GTPase activating effect to promote hydrolysis of GTP by Gαo in rat frontal cortex. FEBS Lett. 1995a;370:135–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous