

Incomplete embryonic lethality and fatal neonatal hemorrhage caused by prothrombin deficiency in mice
- PMID: 9636196
- PMCID: PMC22696
- DOI: 10.1073/pnas.95.13.7603
Incomplete embryonic lethality and fatal neonatal hemorrhage caused by prothrombin deficiency in mice
Abstract
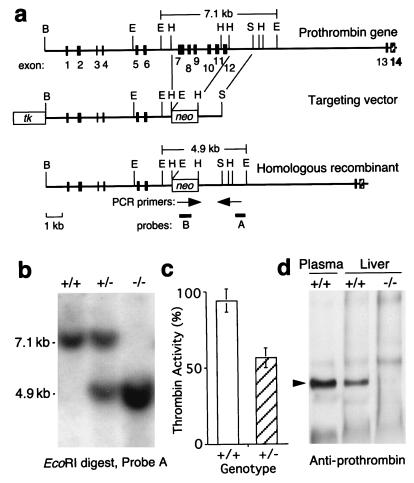
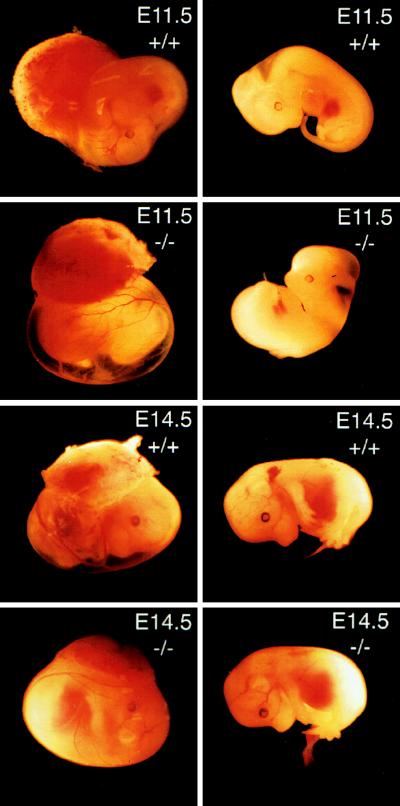
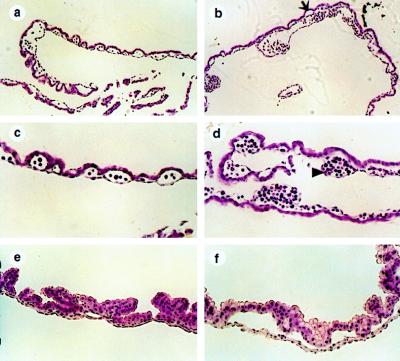
Deficiency of blood coagulation factor V or tissue factor causes the death of mouse embryos by 10.5 days of gestation, suggesting that part of the blood coagulation system is necessary for development. This function is proposed to require either generation of the serine protease thrombin and cell signaling through protease-activated receptors or an activity of tissue factor that is distinct from blood clotting. We find that murine deficiency of prothrombin clotting factor 2 (Cf2) was associated with the death of approximately 50% of Cf2(-/-) embryos by embryonic day 10.5 (E10.5), and surviving embryos had characteristic defects in yolk sac vasculature. Most of the remaining Cf2(-/-) embryos died by E15.5, but those surviving to E18.5 appeared normal. The rare Cf2(-/-) neonates died of hemorrhage on the first postnatal day. These studies suggest that a part of the blood coagulation system is adapted to perform a developmental function. Other mouse models show that the absence of platelets or of fibrinogen does not cause fetal wastage. Therefore, the role of thrombin in development may be independent of its effects on blood coagulation and instead may involve signal transduction on cells other than platelets.
Figures

References
-
- Mann K G. In: Hemostasis and Thrombosis: Basic Principles and Clinical Practice. Colman R W, Hirsch J, Marder V J, Salzman E W, editors. Philadelphia: Lippincott; 1994. pp. 184–199.
-
- Cavanaugh K P, Gurwitz D, Cunningham D D, Bradshaw R A. J Neurochem. 1990;54:1735–1743. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
