

doi: 10.1101/gad.12.12.1775.
Proteasomal regulation of nuclear receptor corepressor-mediated repression
Affiliations
- PMID: 9637679
- PMCID: PMC316907
- DOI: 10.1101/gad.12.12.1775
Item in Clipboard
Proteasomal regulation of nuclear receptor corepressor-mediated repression
Genes Dev.
.
Abstract
Repression of gene transcription is a fundamental property of nuclear hormone receptors. We report here that cell-specific repression by nuclear receptors correlates with levels of nuclear receptor corepressor (N-CoR) protein. N-CoR protein levels are regulated by mSiah2, a mammalian homolog of Drosophila Seven in absentia that targets N-CoR for proteasomal degradation. mSiah2 expression is cell-type specific and differentially regulates the repressive activities of nuclear receptors. These findings establish targeted proteolysis of transcriptional coregulators as a mechanism for cell-specific regulation of gene transcription.
Figures

In vivo and in vitro interactions between N-CoR and mSiah2. (A) Comparison of library clone (mSiah2ΔN), mSiah2, and Sina. The location of conserved Ring finger in mSiah/Sina is shown. (B) Specificity of interactions between N-CoR and mSiah2 in vivo in yeast. SMRT amino acids 1–483 are homologous to N-CoR(1030–1445). (C,D) In vitro interaction studies using GST-pulldown assay. mSiah2(108–325), Sina, and mSin3B (provided by R. Eisenman, Fred Hutchinson Cancer Research Center, Seattle, WA) were 35S-labeled and translated in vitro. The input lane contained 10% of total protein. (E) Coimmunoprecipitation of transfected HA–mSiah2ΔN with N-CoR was performed with nonimmune or rabbit serum containing antibodies raised against N-CoR amino and carboxyl termini. The immunoprecipitated protein was visualized by Western analysis with monoclonal α-HA antibody 12CA5. (F) Mammalian two-hybrid studies. Interaction between cotransfected Gal4–N-CoR(1–160) and VP16–mSiah2ΔN as indicated by activation of a luciferase reporter containing a Gal4 binding site. Gal4 DBD–SMRT(1–483) interacted negligibly with VP16–mSiah2ΔN as shown. Similar results were obtained with Gal4 DBD alone (not shown).
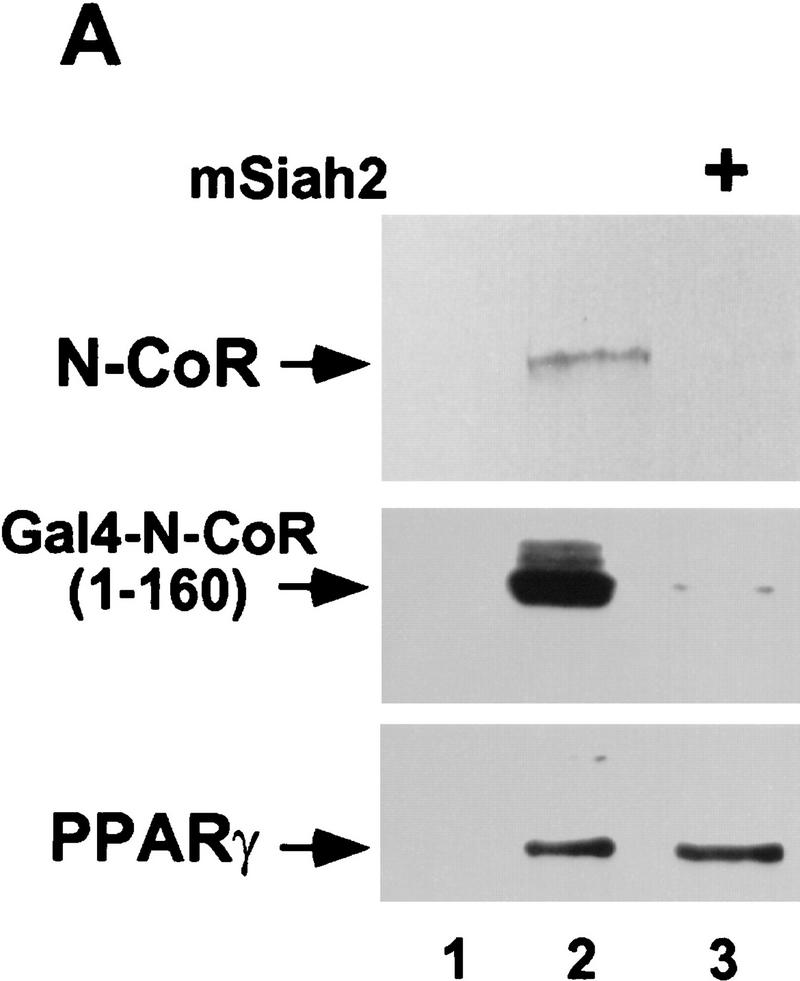
mSiah2 targets N-CoR for proteasomal degradation. (A) mSiah2 blocks expression of N-CoR in 293T cells. Vectors expressing Flag–N-CoR (20 μg, lanes 2,3), Gal4–N-CoR(1–160) (20 μg, lanes 2,3), and PPARγ2 (10 μg, lanes 2,3) or control (lane 1) were transfected into 293T cells with mSiah2 expression vector (lane 3) or control. Gal4–N-CoR and PPARγ were cotransfected. For Flag–N-CoR detection, protein extracts (200 μg) were immunoblotted with monoclonal α-Flag antibody M2. For Gal4–N-CoR and PPARγ detection, 8 μg protein extracts were immunoblotted with either antibody to Gal4 or to PPARγ. (B) Half-life of N-CoR determined by pulse chase experiment. Results were quantitated in a PhosphorImager (Molecular Dynamics), normalized to t = 0 levels, and plotted in semilog format. The level of labeled N-CoR at t = 0 was reduced in mSiah2-transfected cells, attributable to increased degradation of protein synthesized during the time of the pulse. (C) 293T cells were transfected with expression vectors for indicated proteins and either no N-CoR/Gal4–N-CoR(1–160) (lane 1; top, 300 μg protein extract; bottom 29 μg extract), or Flag–N-CoR (300 μg extract, lanes 2–5, top) or Gal4–N-CoR(1–160) (29 μg extract, lanes 2–5, bottom), and treated with or without MG132 (20 μm ). Cell extracts were analyzed by immunoblot analysis.
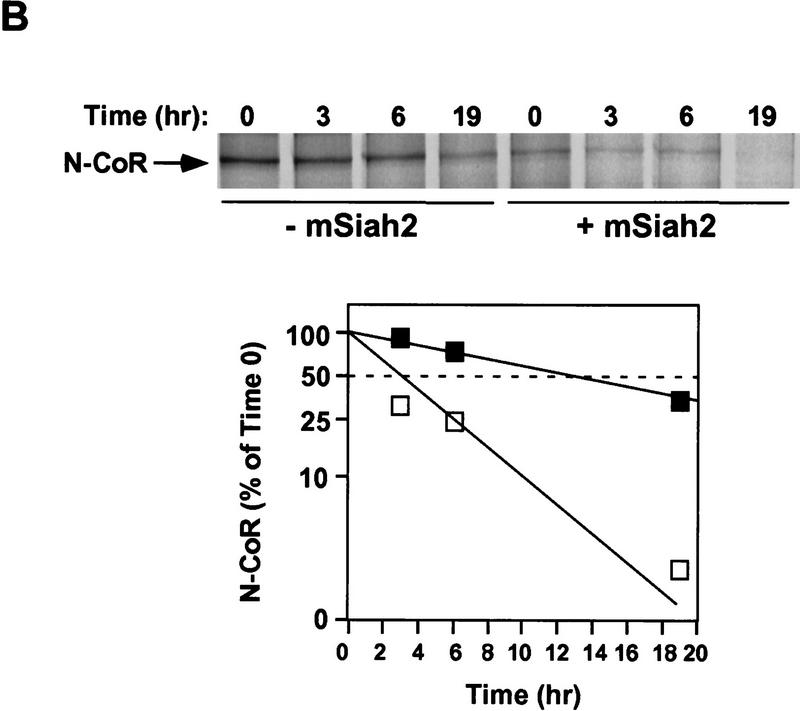
mSiah2 targets N-CoR for proteasomal degradation. (A) mSiah2 blocks expression of N-CoR in 293T cells. Vectors expressing Flag–N-CoR (20 μg, lanes 2,3), Gal4–N-CoR(1–160) (20 μg, lanes 2,3), and PPARγ2 (10 μg, lanes 2,3) or control (lane 1) were transfected into 293T cells with mSiah2 expression vector (lane 3) or control. Gal4–N-CoR and PPARγ were cotransfected. For Flag–N-CoR detection, protein extracts (200 μg) were immunoblotted with monoclonal α-Flag antibody M2. For Gal4–N-CoR and PPARγ detection, 8 μg protein extracts were immunoblotted with either antibody to Gal4 or to PPARγ. (B) Half-life of N-CoR determined by pulse chase experiment. Results were quantitated in a PhosphorImager (Molecular Dynamics), normalized to t = 0 levels, and plotted in semilog format. The level of labeled N-CoR at t = 0 was reduced in mSiah2-transfected cells, attributable to increased degradation of protein synthesized during the time of the pulse. (C) 293T cells were transfected with expression vectors for indicated proteins and either no N-CoR/Gal4–N-CoR(1–160) (lane 1; top, 300 μg protein extract; bottom 29 μg extract), or Flag–N-CoR (300 μg extract, lanes 2–5, top) or Gal4–N-CoR(1–160) (29 μg extract, lanes 2–5, bottom), and treated with or without MG132 (20 μm ). Cell extracts were analyzed by immunoblot analysis.
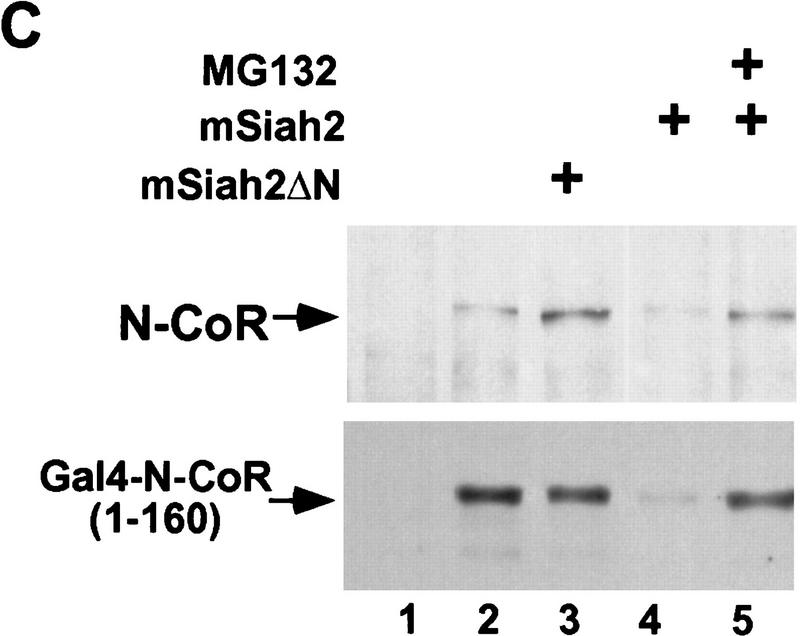
mSiah2 targets N-CoR for proteasomal degradation. (A) mSiah2 blocks expression of N-CoR in 293T cells. Vectors expressing Flag–N-CoR (20 μg, lanes 2,3), Gal4–N-CoR(1–160) (20 μg, lanes 2,3), and PPARγ2 (10 μg, lanes 2,3) or control (lane 1) were transfected into 293T cells with mSiah2 expression vector (lane 3) or control. Gal4–N-CoR and PPARγ were cotransfected. For Flag–N-CoR detection, protein extracts (200 μg) were immunoblotted with monoclonal α-Flag antibody M2. For Gal4–N-CoR and PPARγ detection, 8 μg protein extracts were immunoblotted with either antibody to Gal4 or to PPARγ. (B) Half-life of N-CoR determined by pulse chase experiment. Results were quantitated in a PhosphorImager (Molecular Dynamics), normalized to t = 0 levels, and plotted in semilog format. The level of labeled N-CoR at t = 0 was reduced in mSiah2-transfected cells, attributable to increased degradation of protein synthesized during the time of the pulse. (C) 293T cells were transfected with expression vectors for indicated proteins and either no N-CoR/Gal4–N-CoR(1–160) (lane 1; top, 300 μg protein extract; bottom 29 μg extract), or Flag–N-CoR (300 μg extract, lanes 2–5, top) or Gal4–N-CoR(1–160) (29 μg extract, lanes 2–5, bottom), and treated with or without MG132 (20 μm ). Cell extracts were analyzed by immunoblot analysis.
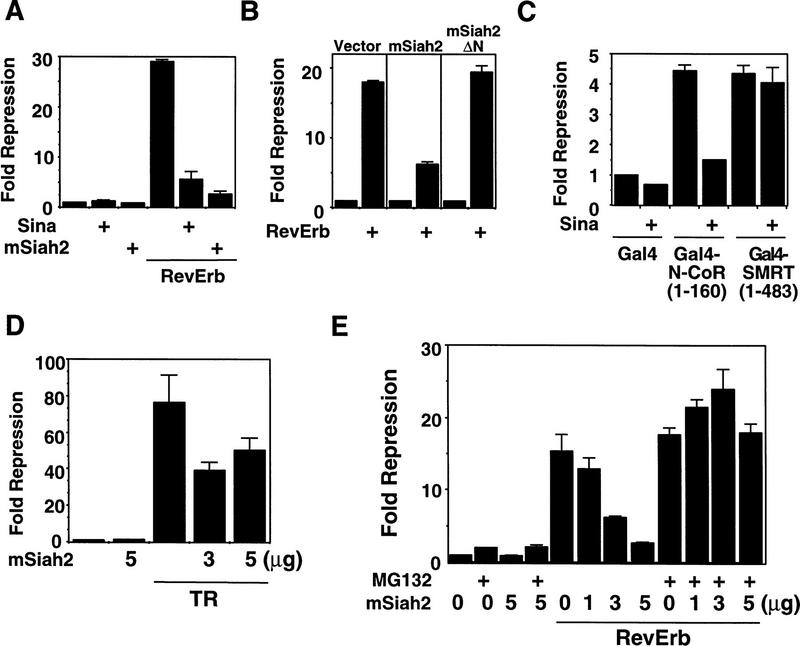
mSiah2 is a nuclear receptor antirepressor. (A) Transfection of mSiah2 and Sina inhibits RevErb repression in 293T cells. Cells were transfected with RevErb expression vector and RevDR2–SV40–luciferase reporter as described previously (Harding and Lazar 1995). (B) mSiah2ΔN does not inhibit RevErb repression in 293T cells. Five micrograms of mSiah2 or mSiah2ΔN expression plasmid was cotransfected where indicated in A and B. (C) Sina also functions as antirepressor for Gal4–N-CoR (1–160) but not Gal4–SMRT(1–483). (D) mSiah2 partially blocks TR repression in 293T cells. Cells were transfected with (Gal4 × 5)–SV40–luciferase and Gal4–TR as described previously (Zhang et al. 1997). (E) MG132 blocks mSiah2 effects on RevErb repression in 293T cells. mSiah2 inhibits RevErb repression in a concentration-dependent manner, and this is prevented by treatment with MG132.
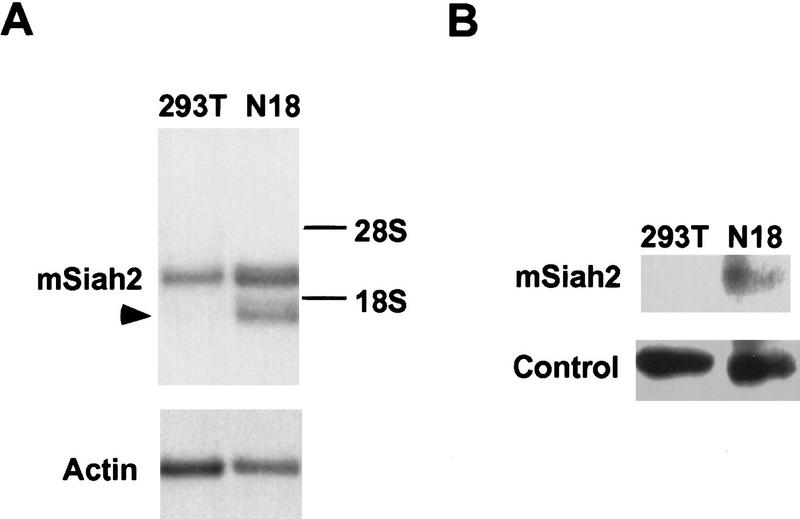
Cell specificity of mSiah2 and N-CoR expression. Northern analysis of mSiah2 (A) and N-CoR (C) mRNA expression. Actin hybridization is shown as loading control. (Arrowhead in A) mSiah2 mRNA species present in N18 cells and absent from 293T cells. Immunoblot analysis of mSiah2 (B) and N-CoR (D) proteins. Total protein (400 μg in B; 200 μg in D) was anlayzed, and equal loading and transfer to nitrocellulose was determined by Ponceau S stain (not shown). Rabbit antiserum used was raised against Sina, but similar results were obtained with antisera of lower titer raised against mSiah2. (B, control) Unidentified protein band (∼42 kD) detected by the Sina antiserum in both N18 and 293T cells. The mSiah2 band comigrated with the in vitro-translated standard (not shown) at ∼36 kD. (D, control) Unidentified protein band (∼215 kD) detected by the N-CoR antiserum in both N18 and 293T cells. The N-CoR band comigrated with in vitro-translated standard (not shown) at ∼270 kD.
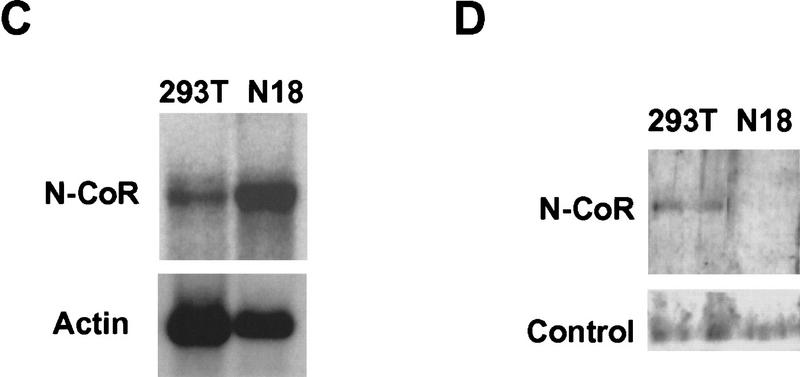
Cell specificity of mSiah2 and N-CoR expression. Northern analysis of mSiah2 (A) and N-CoR (C) mRNA expression. Actin hybridization is shown as loading control. (Arrowhead in A) mSiah2 mRNA species present in N18 cells and absent from 293T cells. Immunoblot analysis of mSiah2 (B) and N-CoR (D) proteins. Total protein (400 μg in B; 200 μg in D) was anlayzed, and equal loading and transfer to nitrocellulose was determined by Ponceau S stain (not shown). Rabbit antiserum used was raised against Sina, but similar results were obtained with antisera of lower titer raised against mSiah2. (B, control) Unidentified protein band (∼42 kD) detected by the Sina antiserum in both N18 and 293T cells. The mSiah2 band comigrated with the in vitro-translated standard (not shown) at ∼36 kD. (D, control) Unidentified protein band (∼215 kD) detected by the N-CoR antiserum in both N18 and 293T cells. The N-CoR band comigrated with in vitro-translated standard (not shown) at ∼270 kD.
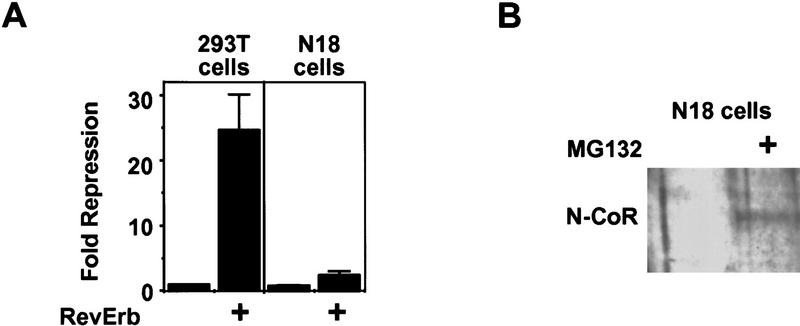
Role of mSiah2-directed proteasomal degradation of N-CoR in nuclear receptor repression in N18 cells. (A) RevErb is a potent repressor in 293T but not in N18 cells. (B) MG132 increases N-CoR protein expression in N18 cells. Western blot performed as in Fig. 4. (C) Effects of MG132 on repression. (Left) MG132 increases RevErb repression in N18 cells; (right) TR represses in N18 cells, and MG132 potentiates TR repression. (D) mSiah2ΔN cotransfection increases RevErb repression in N18 cells. Three micrograms of mSiah2ΔN expression plasmid was transfected.
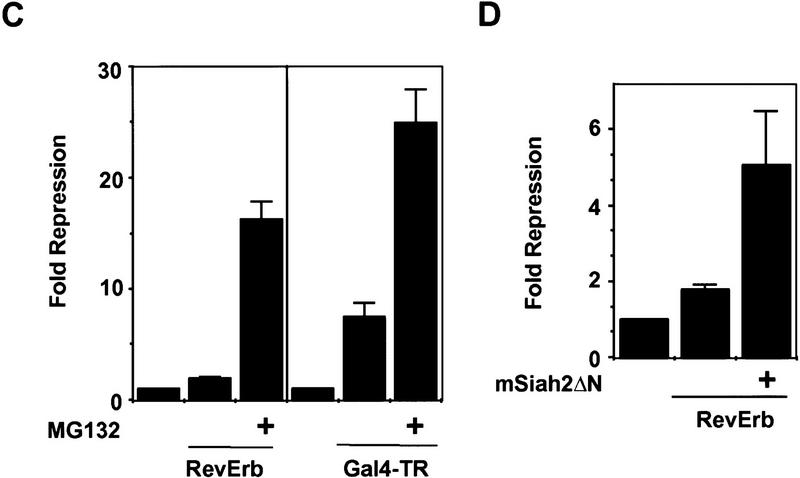
Role of mSiah2-directed proteasomal degradation of N-CoR in nuclear receptor repression in N18 cells. (A) RevErb is a potent repressor in 293T but not in N18 cells. (B) MG132 increases N-CoR protein expression in N18 cells. Western blot performed as in Fig. 4. (C) Effects of MG132 on repression. (Left) MG132 increases RevErb repression in N18 cells; (right) TR represses in N18 cells, and MG132 potentiates TR repression. (D) mSiah2ΔN cotransfection increases RevErb repression in N18 cells. Three micrograms of mSiah2ΔN expression plasmid was transfected.
References
-
- Alland L, Muhle R, Hou H, Potes J, Chin L, Schreiber-Agus N, DePinho RA. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature. 1997;387:49–55. - PubMed
-
- Carthew RW, Rubin GM. seven in absentia, a gene required for specification of R7 cell fate in the Drosophila eye. Cell. 1990;63:561–577. - PubMed
-
- Chang HC, Solomon NM, Wassarman DA, Karim FD, Therrien M, Rubin GM, Wolff T. phyllopod functions in the fate determination of a subset of photoreceptors in Drosophila. Cell. 1995;80:463–472. - PubMed
-
- Chen JD, Evans RM. A transcriptional co-repressor that interacts with nuclear hormone receptors. Nature. 1995;377:454–457. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials