

doi: 10.1128/JB.180.13.3470-3473.1998.
tetA(L) mutants of a tetracycline-sensitive strain of Bacillus subtilis with the polynucleotide phosphorylase gene deleted
Affiliations
- PMID: 9642204
- PMCID: PMC107306
- DOI: 10.1128/JB.180.13.3470-3473.1998
Item in Clipboard
tetA(L) mutants of a tetracycline-sensitive strain of Bacillus subtilis with the polynucleotide phosphorylase gene deleted
J Bacteriol.
1998 Jul.
Abstract
A Bacillus subtilis strain with the polynucleotide phosphorylase gene deleted was sensitive for growth in the presence of tetracycline. This strain was used to select for tetracycline-resistant mutants. A point mutation in the tetA(L) promoter and a spontaneously occurring tetA(L) gene copy number mutant were characterized.
Figures
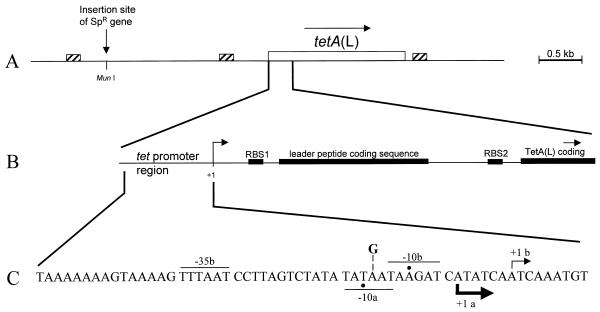
Map of the tetA(L) chromosomal locus. (A) Schematic diagram of the 4.5-kb HindIII fragment cloned into pJBC1 (4). The location of the MunI site, into which the Spr gene was inserted, is shown. Small boxes with diagonal stripes mark the locations of homologous 170-bp direct repeats (1). Open box, tetA(L) gene. (B) Expanded view of the tetA(L) promoter and leader region. Filled boxes, translational signals and coding sequences. The site of transcription initiation is marked +1. RBS, ribosome binding site. (C) Nucleotide sequence of the tetA(L) promoter region. Transcriptional start sites (+1) a and b are indicated, along with the respective −10 and −35 sequences. Actual −10 positions are marked by dots. The thicknesses of the arrows showing the direction of transcription indicate the relative amounts of transcription initiation from the two start sites in the wild type. The location of the A→G change at position −14 (relative to start site b) is shown.
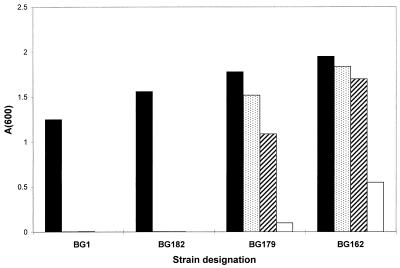
Growth of wild-type and Tcr mutant strains in increasing concentrations of TET. A 2.5-μl portion of an overnight culture was used to inoculate 5 ml of Luria-Bertani broth containing the indicated concentration of TET. Cultures were grown for 16 h with shaking at 37°C, after which the A600 was measured. TET (a 4-mg/ml solution in 50% ethanol) was stored for no more than a few weeks at −10°C. TET concentrations, in micrograms per milliliter, were as follows: 2 (filled bars), 4 (stippled bars), 6 (diagonally striped bars), and 8 (open bars).
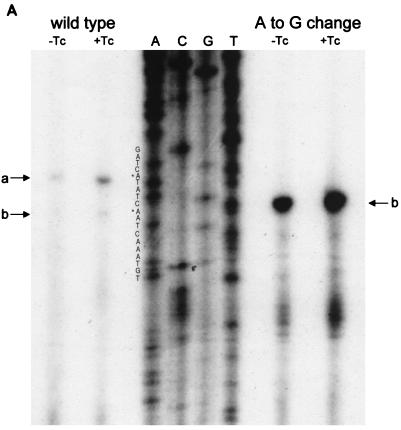
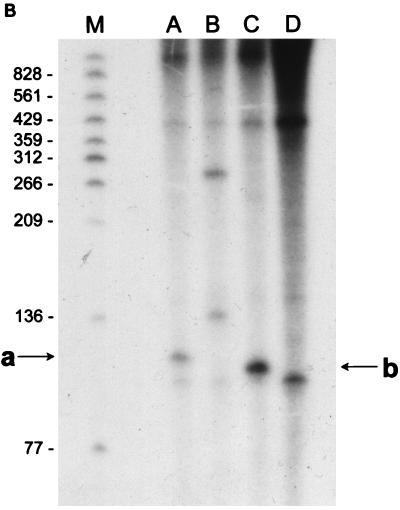
(A) Reverse transcriptase analysis of in vivo tetA(L) mRNA. RNA was isolated from the wild-type strain and from the strain containing the A→G change in the tetA(L) promoter. Strains were grown in the absence or presence of 0.25 μg of TET/ml. Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions. The complement of the ladder sequence, which is the same as the sequence shown in Fig. 1C, is shown on the left of the sequencing ladder. Transcriptional start sites are marked by asterisks. The amount of radioactivity in the bands was quantitated with a PhosphorImager (Molecular Dynamics). (B) In vitro transcription. Plasmid pAN583 contained either a wild-type tetA(L) promoter-leader DNA fragment (lanes A and B), a promoter-leader DNA fragment with the A→G change (lane C), or no tetA(L) DNA (lane D). Plasmid DNA was linearized with XbaI (lanes A, C, and D) or with HindIII (lane B). Digestion with HindIII linearized the plasmid at a site 25 bp downstream of the XbaI site. Comparison of lanes A and B demonstrates that the band indicated by “a” at the left represents transcription from transcriptional start site a. In lane C, the transcription product migrates faster and represents transcription from transcriptional start site b. A nonspecific transcription product runs below band b and is present in all lanes, including lane D [no tetA(L) DNA]. Lane M contained end-labeled DNA fragments from a TaqI restriction endonuclease digest of pSE420 (2). Sizes (in base pairs) of the labeled DNA fragments are indicated on the left. (C) Reverse transcriptase analysis of in vitro-transcribed tetA(L) RNA. Lane 1, wild-type tetA(L) RNA; lane 2, mutant (A→G change) tetA(L) mRNA; lane 3, no RNA (labeled primer only). Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions.
(A) Reverse transcriptase analysis of in vivo tetA(L) mRNA. RNA was isolated from the wild-type strain and from the strain containing the A→G change in the tetA(L) promoter. Strains were grown in the absence or presence of 0.25 μg of TET/ml. Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions. The complement of the ladder sequence, which is the same as the sequence shown in Fig. 1C, is shown on the left of the sequencing ladder. Transcriptional start sites are marked by asterisks. The amount of radioactivity in the bands was quantitated with a PhosphorImager (Molecular Dynamics). (B) In vitro transcription. Plasmid pAN583 contained either a wild-type tetA(L) promoter-leader DNA fragment (lanes A and B), a promoter-leader DNA fragment with the A→G change (lane C), or no tetA(L) DNA (lane D). Plasmid DNA was linearized with XbaI (lanes A, C, and D) or with HindIII (lane B). Digestion with HindIII linearized the plasmid at a site 25 bp downstream of the XbaI site. Comparison of lanes A and B demonstrates that the band indicated by “a” at the left represents transcription from transcriptional start site a. In lane C, the transcription product migrates faster and represents transcription from transcriptional start site b. A nonspecific transcription product runs below band b and is present in all lanes, including lane D [no tetA(L) DNA]. Lane M contained end-labeled DNA fragments from a TaqI restriction endonuclease digest of pSE420 (2). Sizes (in base pairs) of the labeled DNA fragments are indicated on the left. (C) Reverse transcriptase analysis of in vitro-transcribed tetA(L) RNA. Lane 1, wild-type tetA(L) RNA; lane 2, mutant (A→G change) tetA(L) mRNA; lane 3, no RNA (labeled primer only). Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions.
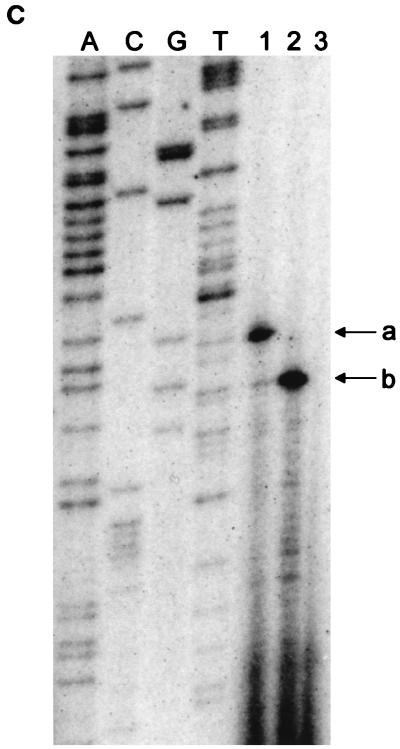
(A) Reverse transcriptase analysis of in vivo tetA(L) mRNA. RNA was isolated from the wild-type strain and from the strain containing the A→G change in the tetA(L) promoter. Strains were grown in the absence or presence of 0.25 μg of TET/ml. Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions. The complement of the ladder sequence, which is the same as the sequence shown in Fig. 1C, is shown on the left of the sequencing ladder. Transcriptional start sites are marked by asterisks. The amount of radioactivity in the bands was quantitated with a PhosphorImager (Molecular Dynamics). (B) In vitro transcription. Plasmid pAN583 contained either a wild-type tetA(L) promoter-leader DNA fragment (lanes A and B), a promoter-leader DNA fragment with the A→G change (lane C), or no tetA(L) DNA (lane D). Plasmid DNA was linearized with XbaI (lanes A, C, and D) or with HindIII (lane B). Digestion with HindIII linearized the plasmid at a site 25 bp downstream of the XbaI site. Comparison of lanes A and B demonstrates that the band indicated by “a” at the left represents transcription from transcriptional start site a. In lane C, the transcription product migrates faster and represents transcription from transcriptional start site b. A nonspecific transcription product runs below band b and is present in all lanes, including lane D [no tetA(L) DNA]. Lane M contained end-labeled DNA fragments from a TaqI restriction endonuclease digest of pSE420 (2). Sizes (in base pairs) of the labeled DNA fragments are indicated on the left. (C) Reverse transcriptase analysis of in vitro-transcribed tetA(L) RNA. Lane 1, wild-type tetA(L) RNA; lane 2, mutant (A→G change) tetA(L) mRNA; lane 3, no RNA (labeled primer only). Transcriptional start sites are marked a and b, as in Fig. 1. Lanes A, C, G, and T are a sequence ladder obtained with the primer used for the reverse transcriptase reactions.
References
-
- Amano H, Shishido K. Bacillus subtilis strains carry highly homologous direct repeat sequences on their chromosomes. Biosci Biotechnol Biochem. 1995;59:2149–2150. - PubMed
-
- Brosius J. Compilation of superlinker vectors. Methods Enzymol. 1992;216:469–483. - PubMed
-
- Cheng J, Guffanti A A, Krulwich T A. The chromosomal tetracycline-resistance locus of Bacillus subtilis encodes a Na+/H+ antiporter that is physiologically important at elevated pH. J Biol Chem. 1994;269:27365–27371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
