

Localization, dynamics, and protein interactions reveal distinct roles for ER and Golgi SNAREs
- PMID: 9647643
- PMCID: PMC2133002
- DOI: 10.1083/jcb.141.7.1489
Localization, dynamics, and protein interactions reveal distinct roles for ER and Golgi SNAREs
Erratum in
- J Cell Biol 1998 Aug 10;142(3):following 881
Abstract
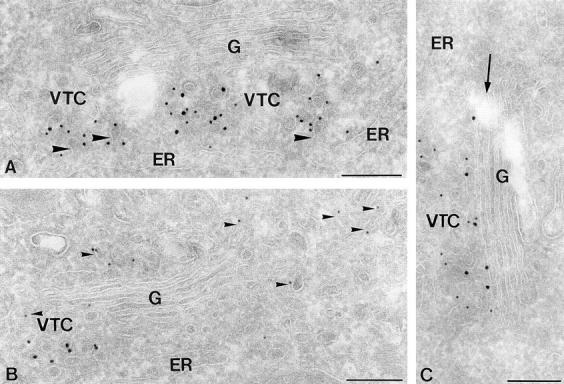
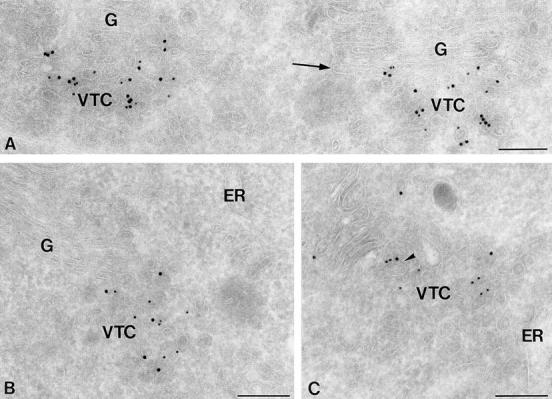
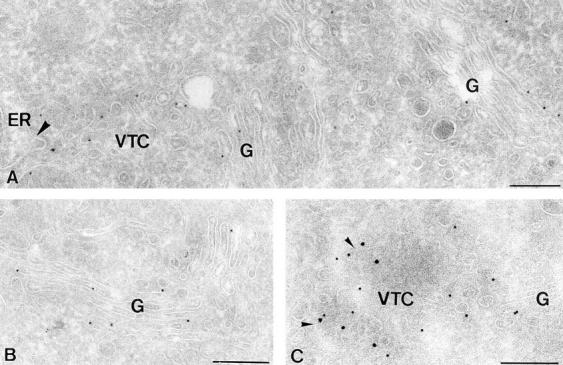
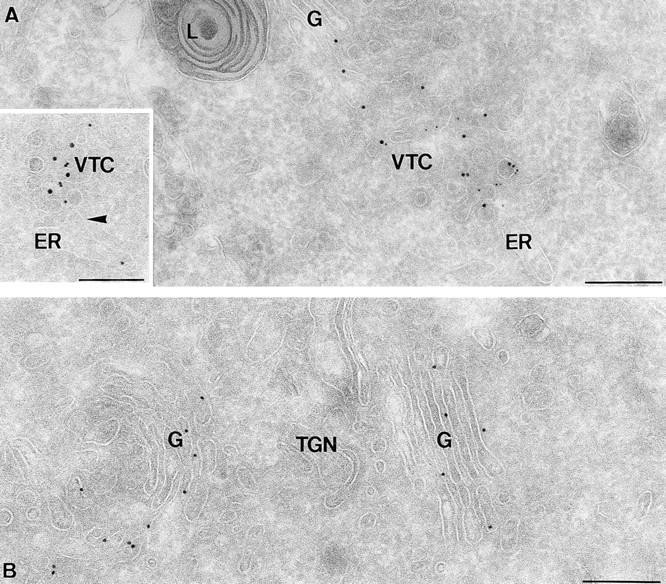
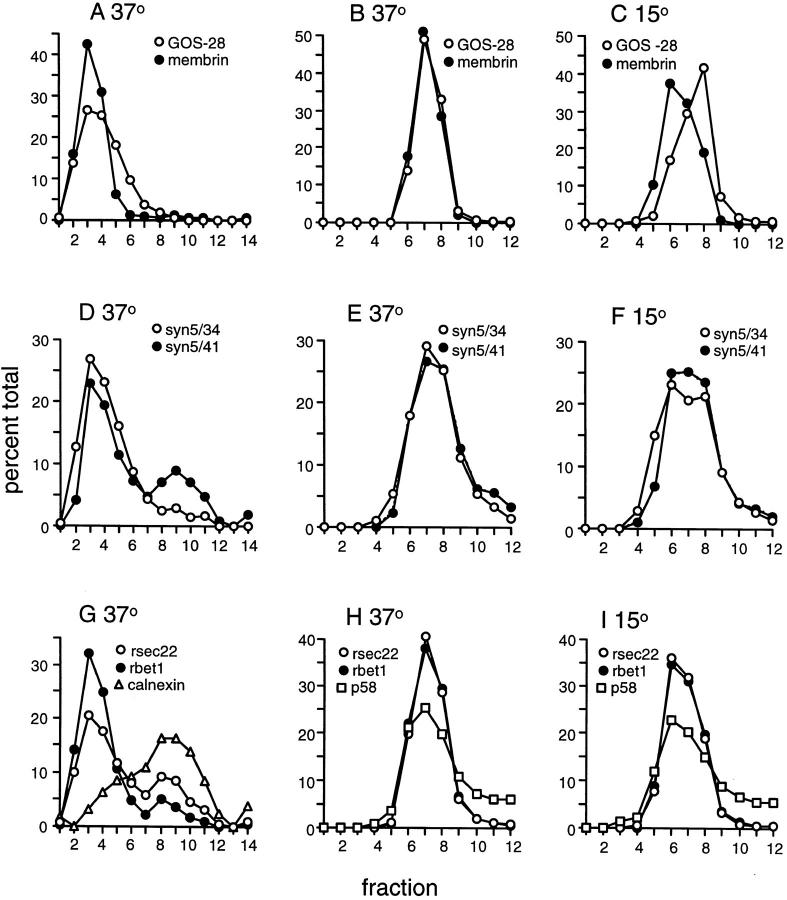
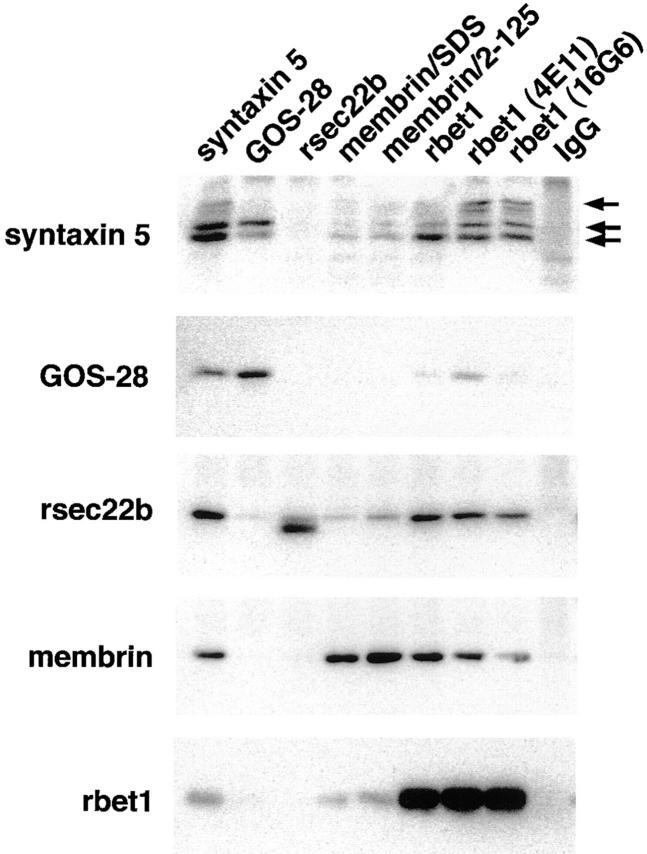
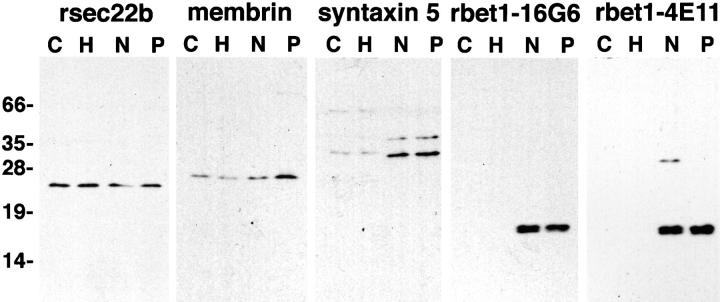
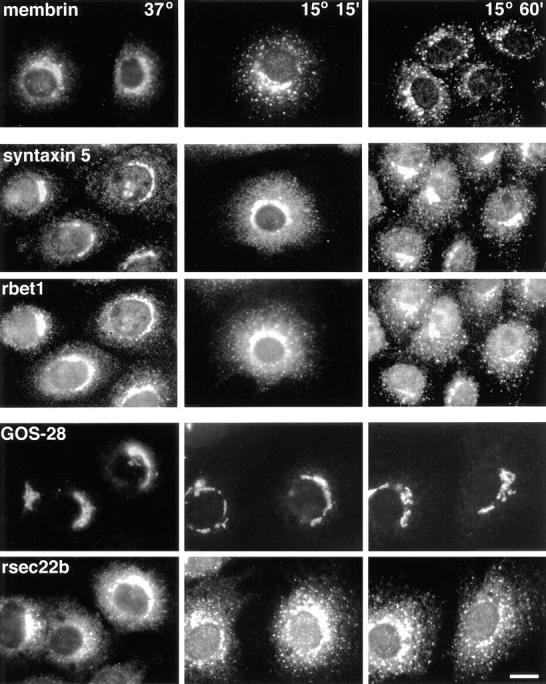
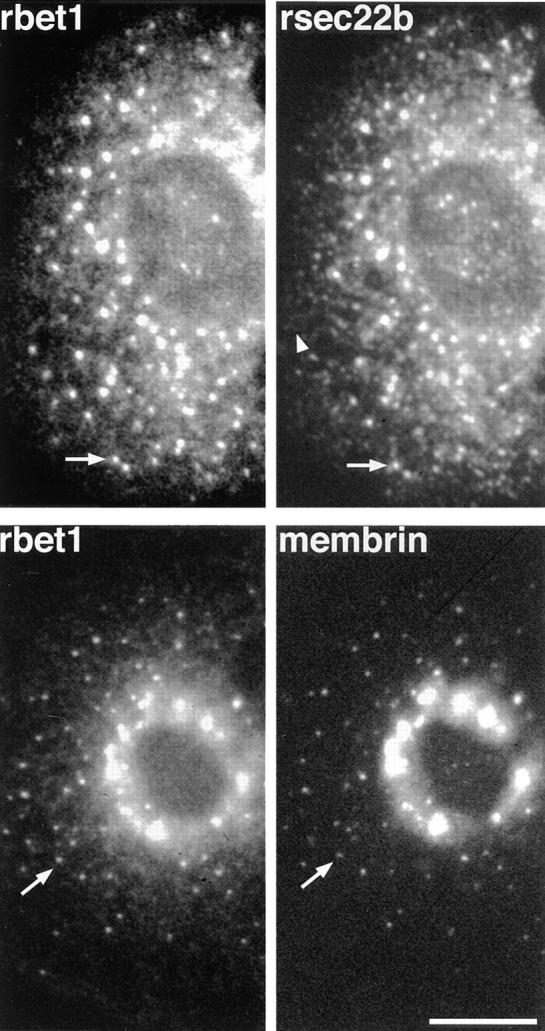
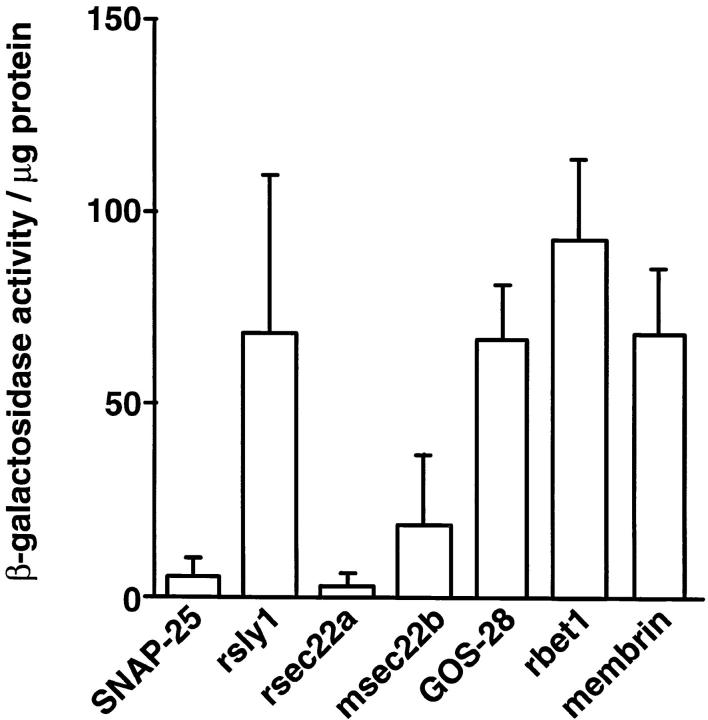
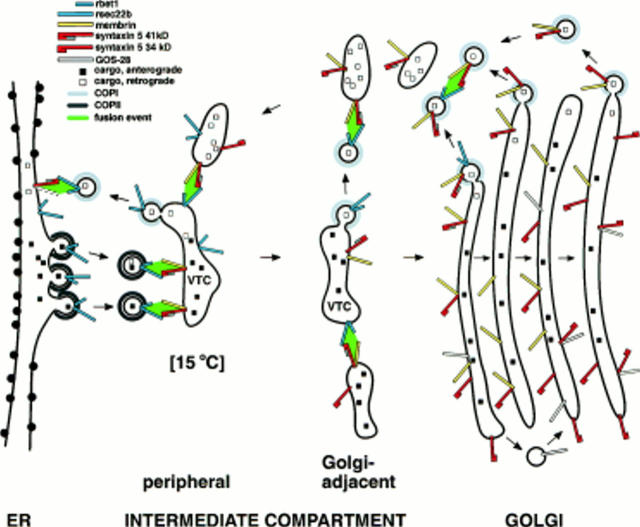
ER-to-Golgi transport, and perhaps intraGolgi transport involves a set of interacting soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) proteins including syntaxin 5, GOS-28, membrin, rsec22b, and rbet1. By immunoelectron microscopy we find that rsec22b and rbet1 are enriched in COPII-coated vesicles that bud from the ER and presumably fuse with nearby vesicular tubular clusters (VTCs). However, all of the SNAREs were found on both COPII- and COPI-coated membranes, indicating that similar SNARE machinery directs both vesicle pathways. rsec22b and rbet1 do not appear beyond the first Golgi cisterna, whereas syntaxin 5 and membrin penetrate deeply into the Golgi stacks. Temperature shifts reveal that membrin, rsec22b, rbet1, and syntaxin 5 are present together on membranes that rapidly recycle between peripheral and Golgi-centric locations. GOS-28, on the other hand, maintains a fixed localization in the Golgi. By immunoprecipitation analysis, syntaxin 5 exists in at least two major subcomplexes: one containing syntaxin 5 (34-kD isoform) and GOS-28, and another containing syntaxin 5 (41- and 34-kD isoforms), membrin, rsec22b, and rbet1. Both subcomplexes appear to involve direct interactions of each SNARE with syntaxin 5. Our results indicate a central role for complexes among rbet1, rsec22b, membrin, and syntaxin 5 (34 and 41 kD) at two membrane fusion interfaces: the fusion of ER-derived vesicles with VTCs, and the assembly of VTCs to form cis-Golgi elements. The 34-kD syntaxin 5 isoform, membrin, and GOS-28 may function in intraGolgi transport.
Figures
References
-
- Bennett MK, Garcia-Arraras JE, Elferink LA, Peterson K, Fleming AM, Hazuka CD, Scheller RH. The syntaxin family of vesicular transport receptors. Cell. 1993;74:863–873. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
