

Molecular dissection of the Rho-associated protein kinase (p160ROCK)-regulated neurite remodeling in neuroblastoma N1E-115 cells
- PMID: 9647654
- PMCID: PMC2133015
- DOI: 10.1083/jcb.141.7.1625
Molecular dissection of the Rho-associated protein kinase (p160ROCK)-regulated neurite remodeling in neuroblastoma N1E-115 cells
Abstract
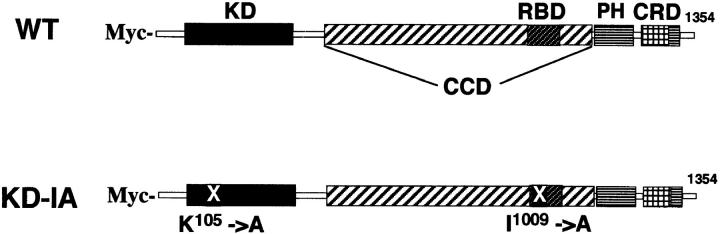
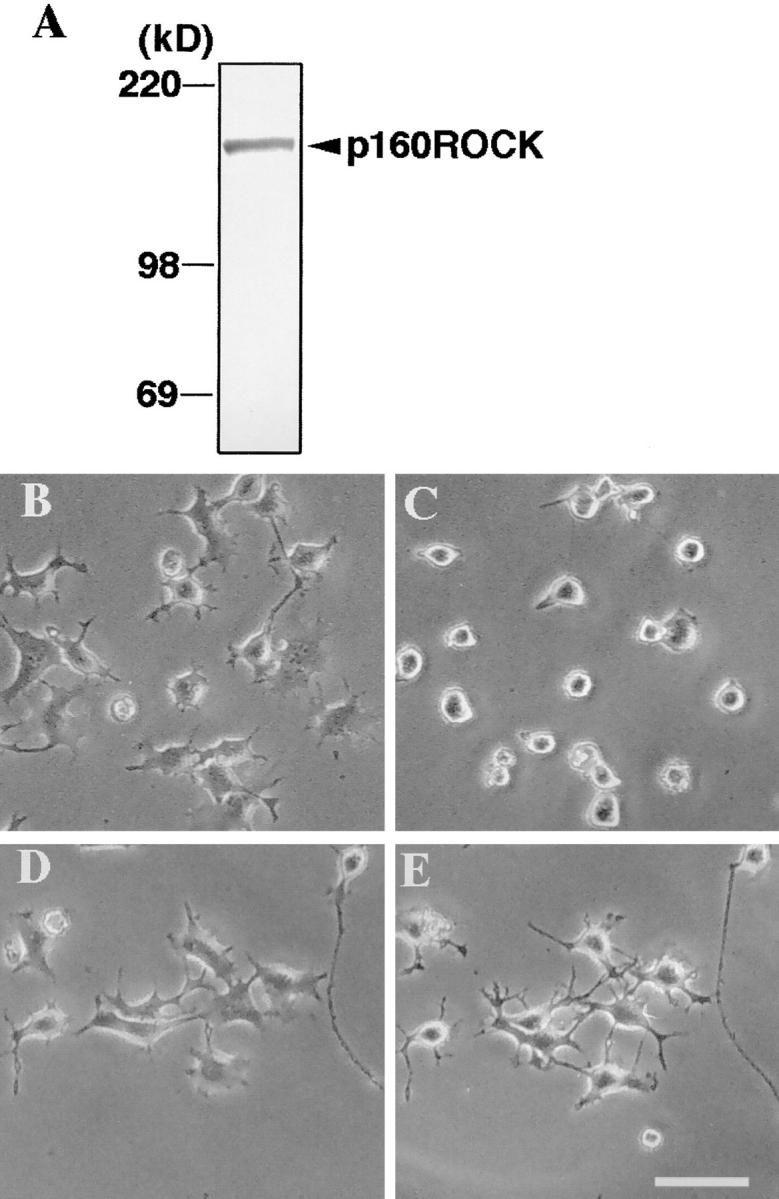
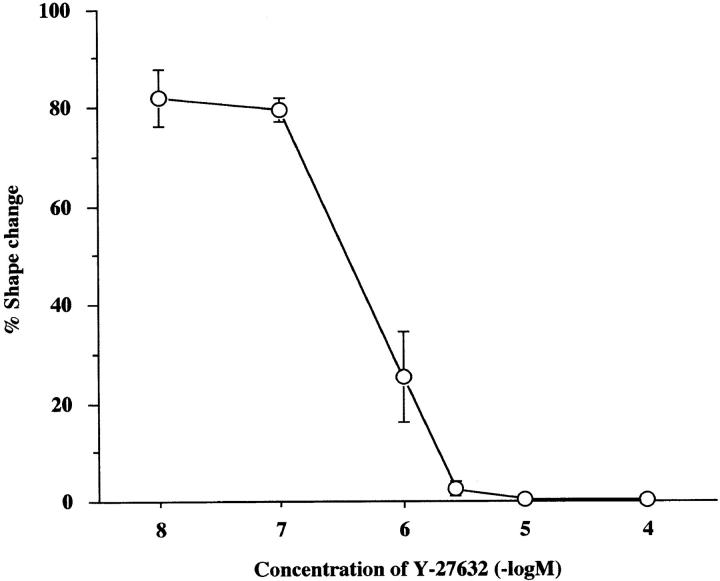
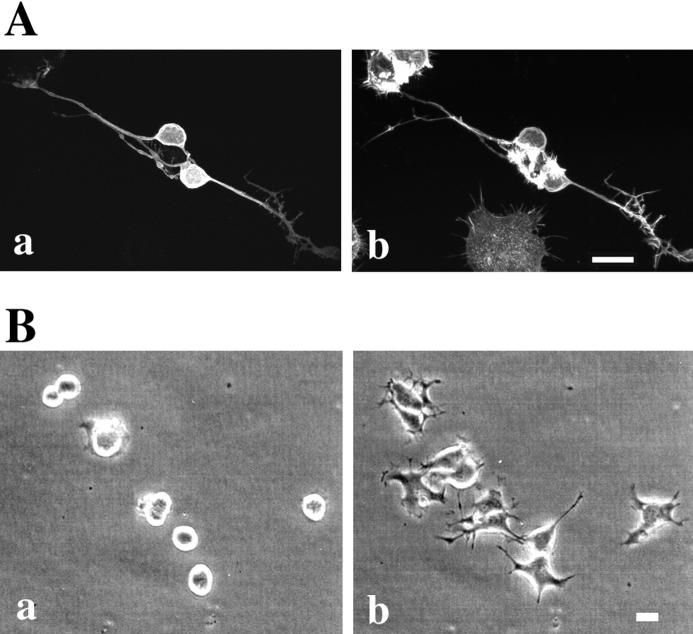
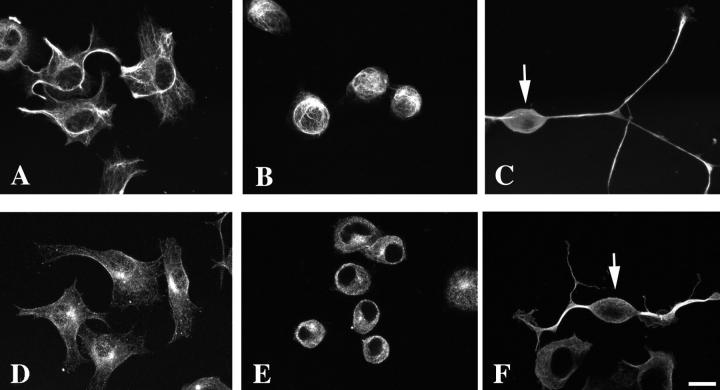
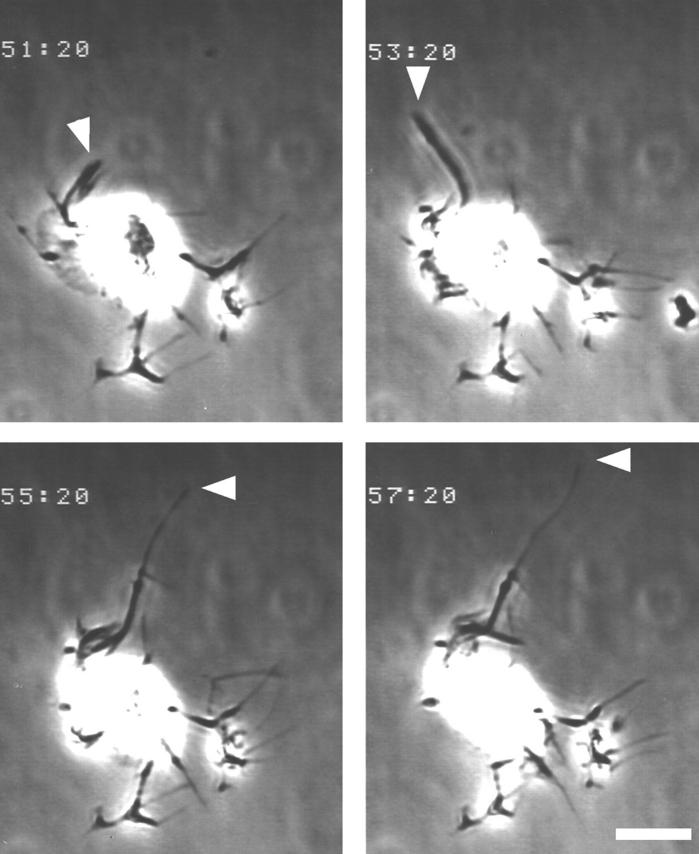
A critical role for the small GTPase Rho and one of its targets, p160ROCK (a Rho-associated coiled coil-forming protein kinase), in neurite remodeling was examined in neuroblastoma N1E-115 cells. Using wild-type and a dominant-negative form of p160ROCK and a p160ROCK-specific inhibitor, Y-27632, we show here that p160ROCK activation is necessary and sufficient for the agonist-induced neurite retraction and cell rounding. The neurite retraction was accompanied by elevated phosphorylation of myosin light chain and the disassembly of the intermediate filaments and microtubules. Y-27632 blocked both neurite retraction and the elevation of myosin light chain phosphorylation in a similar concentration-dependent manner. On the other hand, suppression of p160ROCK activity by expression of a dominant-negative form of p160ROCK induced neurites in the presence of serum by inducing the reassembly of the intermediate filaments and microtubules. The neurite outgrowth by the p160ROCK inhibition was blocked by coexpression of dominant-negative forms of Cdc42 and Rac, indicating that p160ROCK constitutively and negatively regulates neurite formation at least in part by inhibiting activation of Cdc42 and Rac. The assembly of microtubules and intermediate filaments to form extended processes by inhibitors of the Rho-ROCK pathway was also observed in Swiss 3T3 cells. These results indicate that Rho/ROCK-dependent tonic inhibition of cell process extension is exerted via activation of the actomysin-based contractility, in conjunction with a suppression of assembly of intermediate filaments and microtubules in many cell types including, but not exclusive to, neuronal cells.
Figures







References
-
- Amano M, Itoh M, Kimura K, Fukata Y, Chihara K, Nakano T, Mastuura Y, Kaibuchi K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase) J Biol Chem. 1996;271:20246–20249. - PubMed
-
- Amano M, Chihara K, Kimura K, Fukata Y, Nakamura N, Mastuura Y, Kaibuchi K. Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science. 1997;275:1308–1311. - PubMed
-
- Aletta JM, Shelanski ML, Greene LA. Phosphorylation of the peripherin 58-kD, a neuronal intermediate filament protein. Regulation by nerve growth factor and other agents. J Biol Chem. 1989;264:4619–4627. - PubMed
-
- Bently D, O'Connor TP. Cytoskeletal events in growth cone steering. Curr Opin Neurobiol. 1994;4:43–48. - PubMed
-
- Daniels M. The role of microtubules in the growth and stabilization of nerve fibers. Annu NY Acad Sci. 1975;253:535–544. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
