

Complementary roles for receptor clustering and conformational change in the adhesive and signaling functions of integrin alphaIIb beta3
- PMID: 9647659
- PMCID: PMC2133003
- DOI: 10.1083/jcb.141.7.1685
Complementary roles for receptor clustering and conformational change in the adhesive and signaling functions of integrin alphaIIb beta3
Abstract
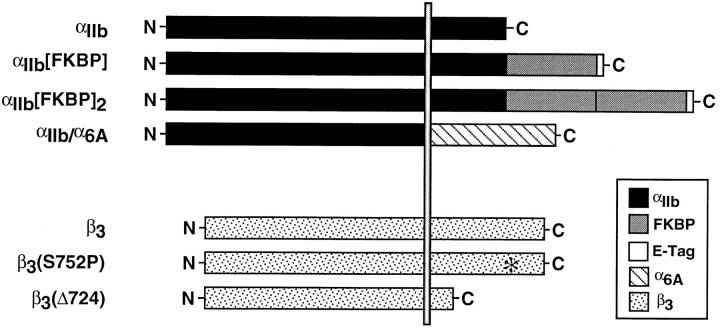
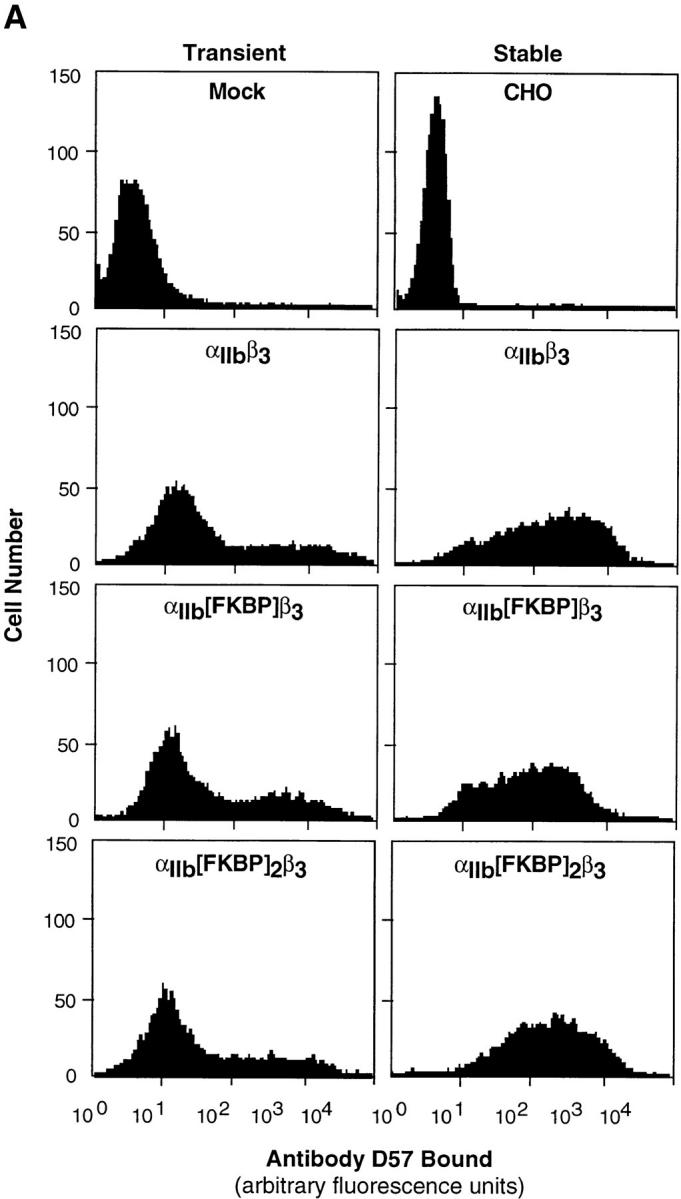
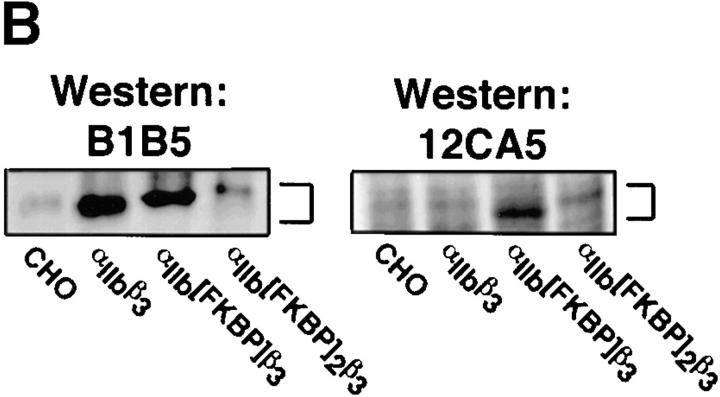
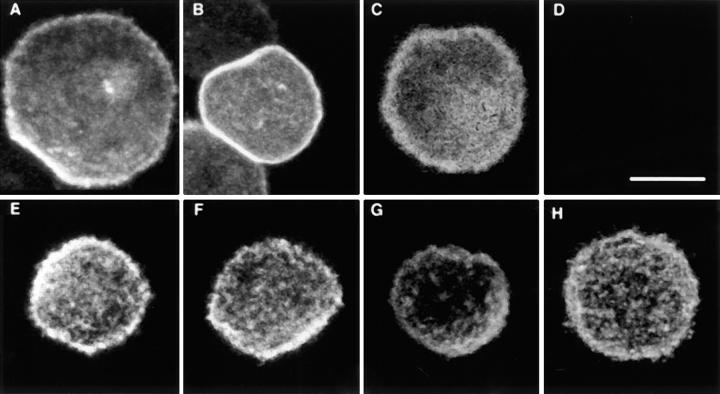
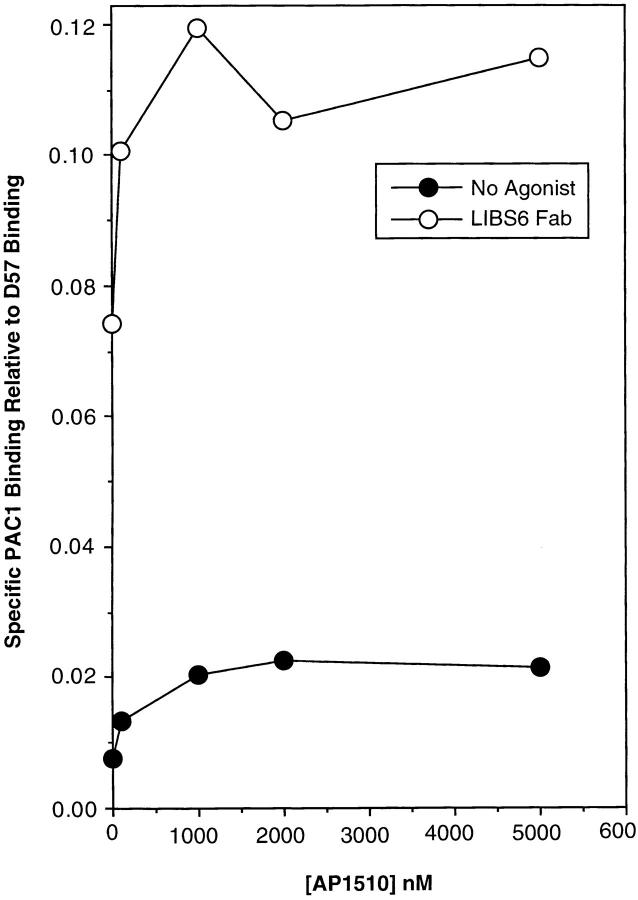
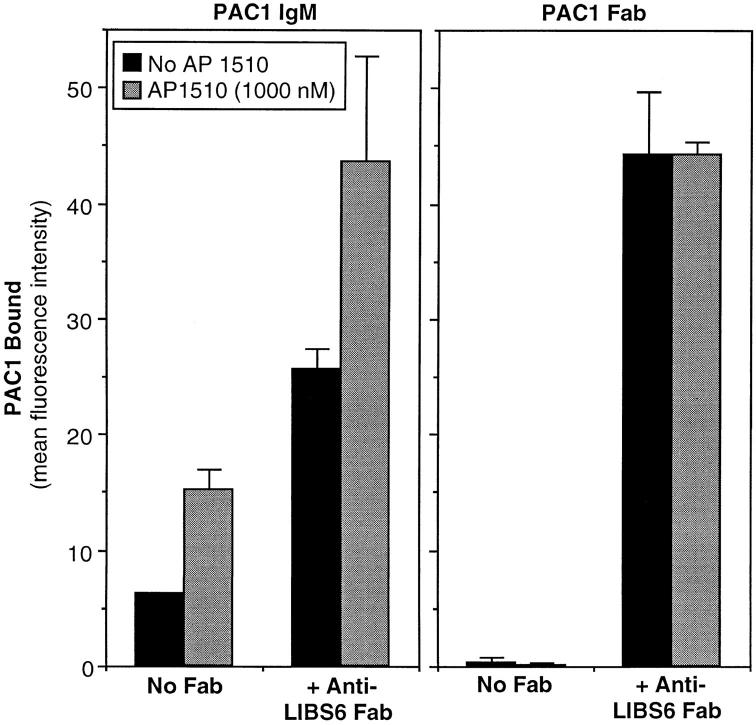
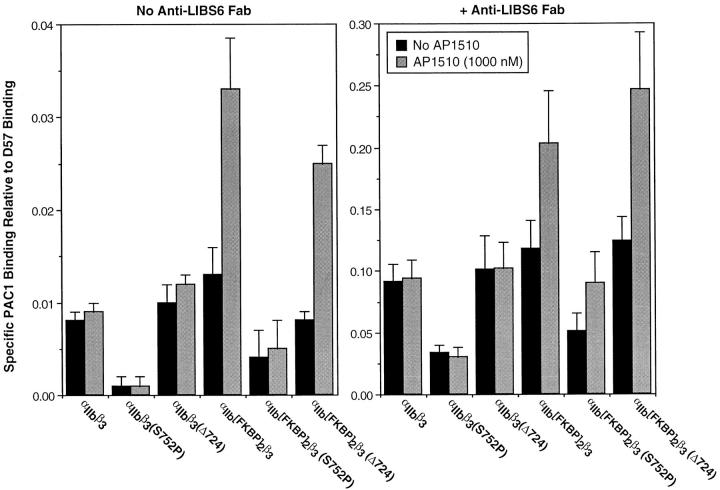
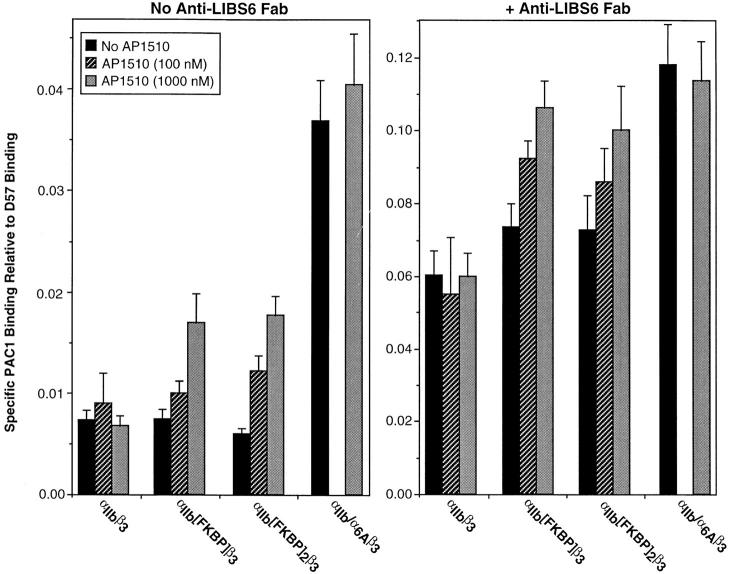
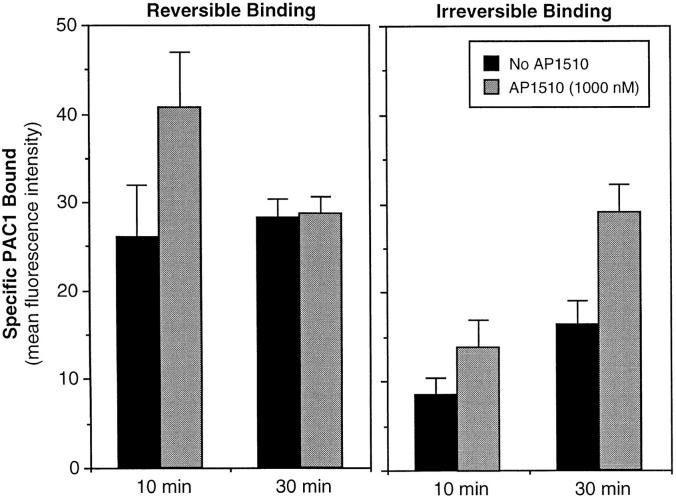
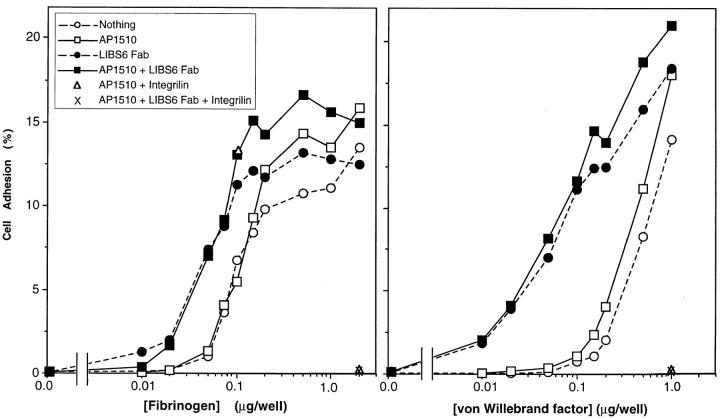
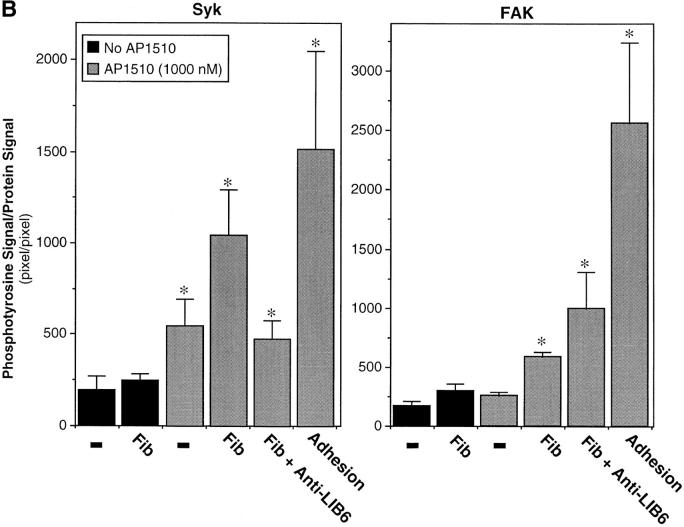
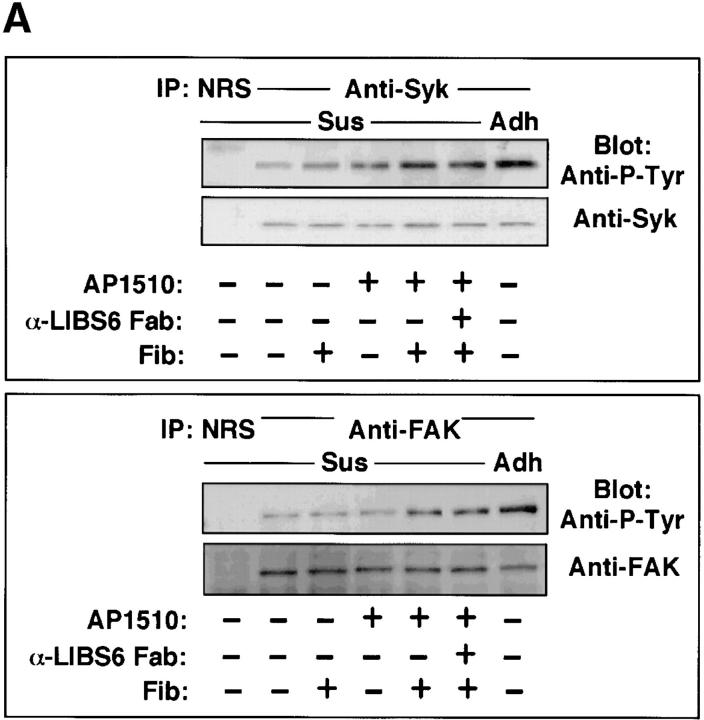
Integrin alphaIIb beta3 mediates platelet aggregation and "outside-in" signaling. It is regulated by changes in receptor conformation and affinity and/or by lateral diffusion and receptor clustering. To document the relative contributions of conformation and clustering to alphaIIb beta3 function, alphaIIb was fused at its cytoplasmic tail to one or two FKBP12 repeats (FKBP). These modified alphaIIb subunits were expressed with beta3 in CHO cells, and the heterodimers could be clustered into morphologically detectable oligomers upon addition of AP1510, a membrane-permeable, bivalent FKBP ligand. Integrin clustering by AP1510 caused binding of fibrinogen and a multivalent (but not monovalent) fibrinogen-mimetic antibody. However, ligand binding due to clustering was only 25-50% of that observed when alphaIIb beta3 affinity was increased by an activating antibody or an activating mutation. The effects of integrin clustering and affinity modulation were additive, and clustering promoted irreversible ligand binding. Clustering of alphaIIb beta3 also promoted cell adhesion to fibrinogen or von Willebrand factor, but not as effectively as affinity modulation. However, clustering was sufficient to trigger fibrinogen-independent tyrosine phosphorylation of pp72(Syk) and fibrinogen-dependent phosphorylation of pp125(FAK), even in non-adherent cells. Thus, receptor clustering and affinity modulation play complementary roles in alphaIIb beta3 function. Affinity modulation is the predominant regulator of ligand binding and cell adhesion, but clustering increases these responses further and triggers protein tyrosine phosphorylation, even in the absence of affinity modulation. Both affinity modulation and clustering may be needed for optimal function of alphaIIb beta3 in platelets.
Figures
References
-
- Abrams C, Deng J, Steiner B, Shattil SJ. Determinants of specificity of a baculovirus-expressed antibody Fab fragment that binds selectively to the activated form of integrin αIIbβ3 . J Biol Chem. 1994;269:18781–18788. - PubMed
-
- Abrams CS, Ruggeri ZM, Taub R, Hoxie JA, Nagaswami C, Weisel W, Shattil SJ. Anti-idiotypic antibodies against an antibody to the platelet glycoprotein (GP) IIb-IIIa complex mimic GP IIb-IIIa by recognizing fibrinogen. J Biol Chem. 1992;267:2775–2785. - PubMed
-
- Banfic H, Tang XW, Batty IH, Downes CP, Chen CS, Rittenhouse SE. A novel integrin-activated pathway forms PKB/Akt-stimulatory phosphatidylinositol 3,4-bisphosphate via phosphatidylinositol 3-phosphate in platelets. J Biol Chem. 1998;273:13–16. - PubMed
-
- Bazzoni G, Hemler ME. Are changes in integrin affinity and conformation overemphasized? . Trends Biochem Sci. 1998;23:30–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
