

Proneural gene self-stimulation in neural precursors: an essential mechanism for sense organ development that is regulated by Notch signaling
- PMID: 9649507
- PMCID: PMC316959
- DOI: 10.1101/gad.12.13.2036
Proneural gene self-stimulation in neural precursors: an essential mechanism for sense organ development that is regulated by Notch signaling
Abstract
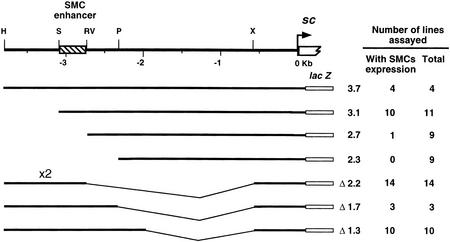
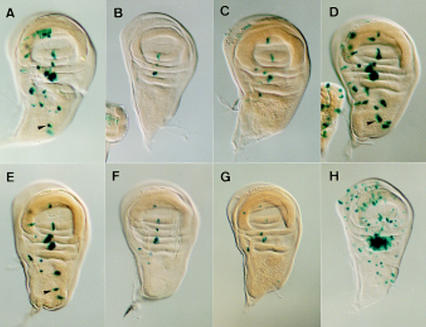
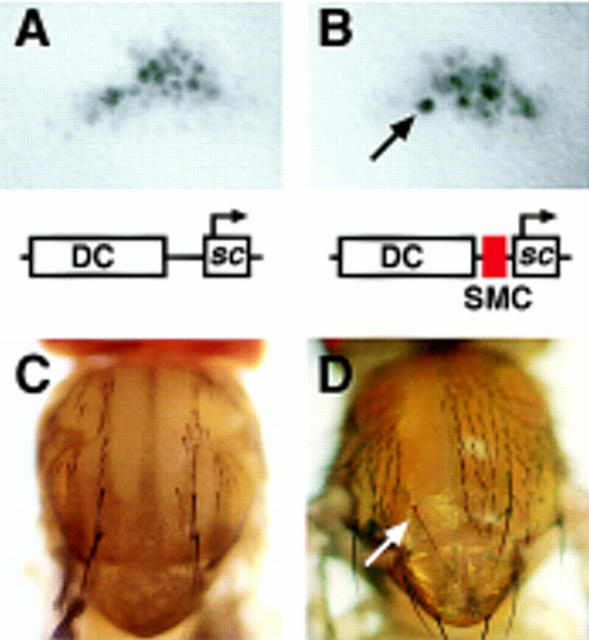
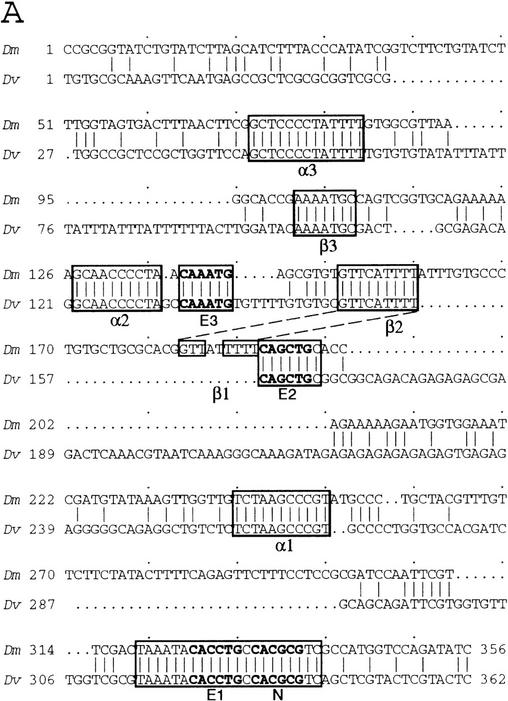
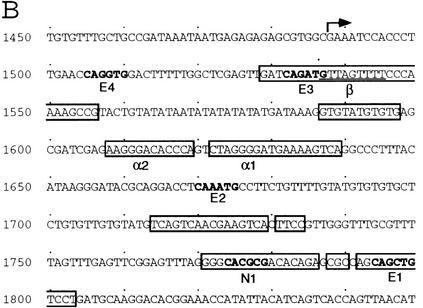
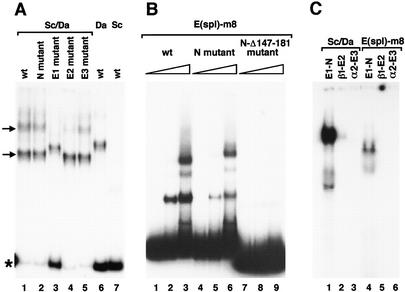
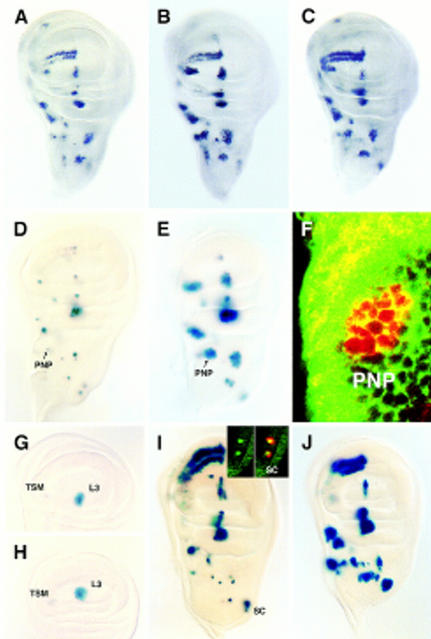
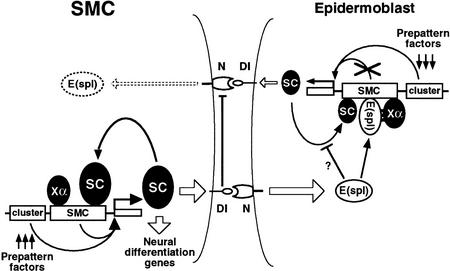
To learn about the acquisition of neural fate by ectodermal cells, we have analyzed a very early sign of neural commitment in Drosophila, namely the specific accumulation of achaete-scute complex (AS-C) proneural proteins in the cell that becomes a sensory organ mother cell (SMC). We have characterized an AS-C enhancer that directs expression specifically in SMCs. This enhancer promotes Scute protein accumulation in these cells, an event essential for sensory organ development in the absence of other AS-C genes. Interspecific sequence comparisons and site-directed mutagenesis show the presence of several conserved motifs necessary for enhancer action, some of them binding sites for proneural proteins. These and other data indicate that the enhancer mediates scute self-stimulation, although only in the presence of additional activating factors, which most likely interact with conserved motifs reminiscent of NF-kappaB-binding sites. Cells neighboring the SMC do not acquire the neural fate because the Notch signaling pathway effectors, the Enhancer of split bHLH proteins, block this proneural gene self-stimulatory loop, possibly by antagonizing the action on the enhancer of the NF-kappaB-like factors or the proneural proteins. These data suggest a mechanism for SMC committment.
Figures

References
-
- Arora K, Dai H, Kazuko SG, Jamal J, O’Connor MB, Letsou A, Warrior R. The Drosophila schnurri gene acts in the Dpp/TGFβ signalling pathway and encodes a transcription factor homologous to the human MBP family. Cell. 1995;81:781–790. - PubMed
-
- Artavanis-Tsakonas S, Simpson P. Choosing a cell fate: A view from the Notch locus. Trends Genet. 1991;7:403–408. - PubMed
-
- Bailey AM, Posakony JW. Suppressor of Hairless directly activates transcription of Enhancer of split complex genes in response to Notch receptor activity. Genes & Dev. 1995;9:2609–2622. - PubMed
-
- Bang AG, Bailey AM, Posakony JW. Hairless promotes stable commitment to the sensory organ precursor cell fate by negatively regulating the activity of the Notch signaling pathway. Dev Biol. 1995;172:479–494. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous