

The neural cell adhesion molecule L1 interacts with the AP-2 adaptor and is endocytosed via the clathrin-mediated pathway
- PMID: 9651214
- PMCID: PMC1226881
- DOI: 10.1523/JNEUROSCI.18-14-05311.1998
The neural cell adhesion molecule L1 interacts with the AP-2 adaptor and is endocytosed via the clathrin-mediated pathway
Abstract
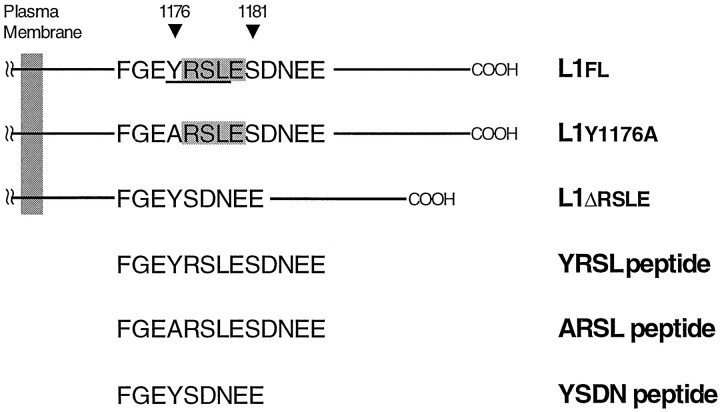
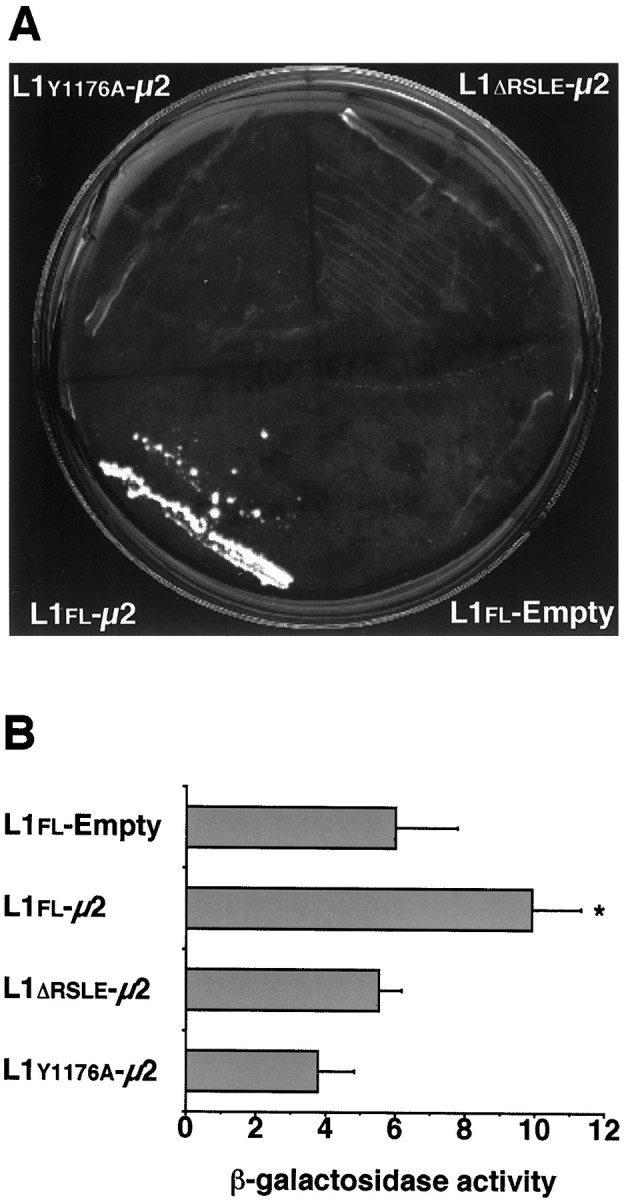
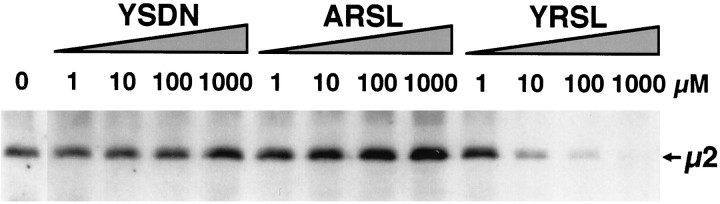
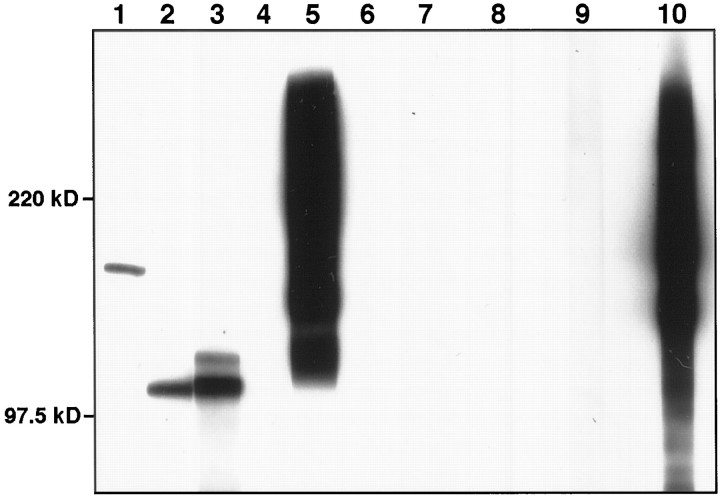
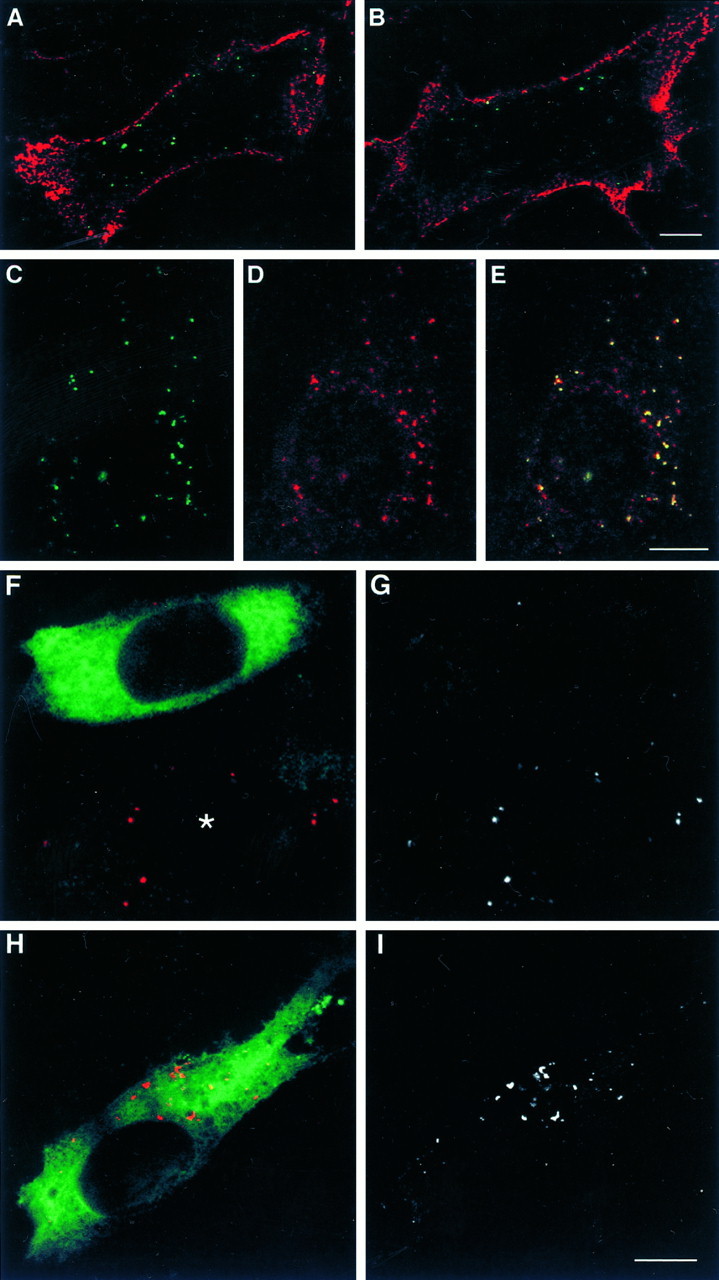
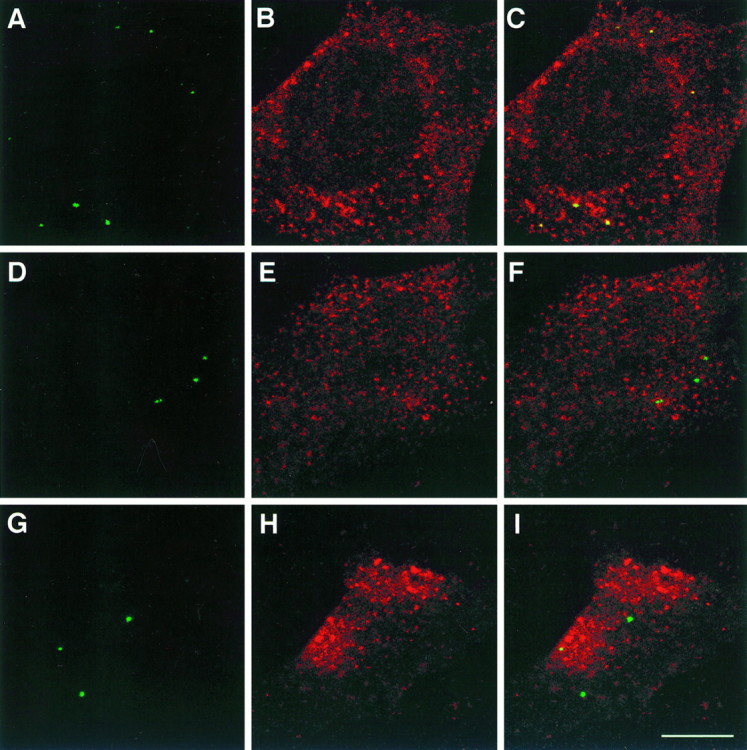
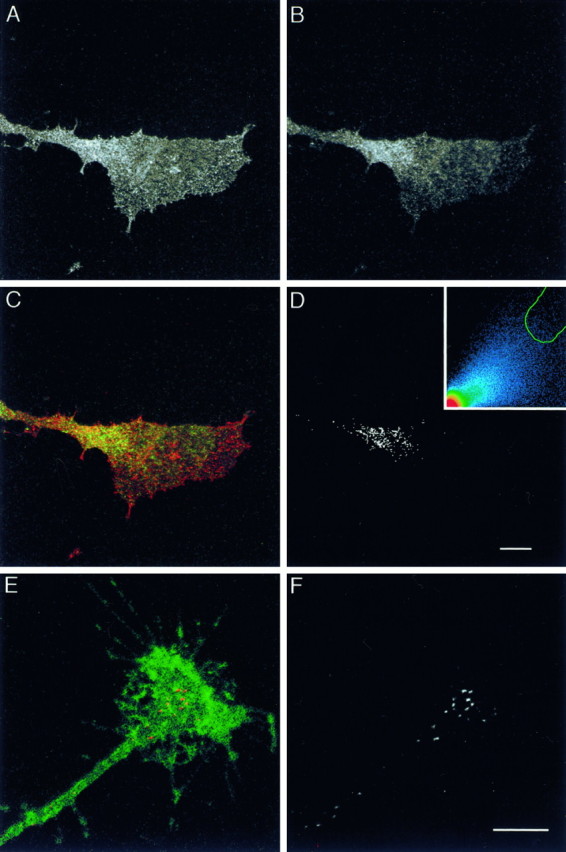
Cell-cell interactions mediated via cell adhesion molecules (CAMs) are dynamically regulated during nervous system development. One mechanism to control the amount of cell surface CAMs is to regulate their recycling from the plasma membrane. The L1 subfamily of CAMs has a highly conserved cytoplasmic domain that contains a tyrosine, followed by the alternatively spliced RSLE (Arg-Ser-Leu-Glu) sequence. The resulting sequence of YRSL conforms to a tyrosine-based sorting signal that mediates clathrin-dependent endocytosis of signal-bearing proteins. The present study shows that L1 associates in rat brain with AP-2, a clathrin adaptor that captures plasma membrane proteins with tyrosine-based signals for endocytosis by coated pits. In vitro assays demonstrate that this interaction occurs via the YRSL sequence of L1 and the mu 2 chain of AP-2. In L1-transfected 3T3 cells, L1 endocytosis is blocked by dominant-negative dynamin that specifically disrupts clathrin-mediated internalization. Furthermore, endocytosed L1 colocalizes with the transferrin receptor (TfR), a marker for clathrin-mediated internalization. Mutant forms of L1 that lack the YRSL do not colocalize with TfR, indicating that the YRSL mediates endocytosis of L1. In neurons, L1 is endocytosed preferentially at the rear of axonal growth cones, colocalizing with Eps15, another marker for the clathrin endocytic pathway. These results establish a mechanism by which L1 can be internalized from the cell surface and suggest that an active region of L1 endocytosis at the rear of growth cones is important in L1-dependent axon growth.
Figures
References
-
- Aberle H, Schwartz H, Kemler R. Cadherin–catenin complex: protein interactions and their implications for cadherin function. J Cell Biochem. 1996;61:514–523. - PubMed
-
- Bailey CH, Chen M, Keller F, Kandel ER. Serotonin-mediated endocytosis of apCAM: an early step of learning-related synaptic growth in Aplysia. Science. 1992;256:645–649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous