

CD4 T cell tolerance to human C-reactive protein, an inducible serum protein, is mediated by medullary thymic epithelium
- PMID: 9653079
- PMCID: PMC2525550
- DOI: 10.1084/jem.188.1.5
CD4 T cell tolerance to human C-reactive protein, an inducible serum protein, is mediated by medullary thymic epithelium
Abstract
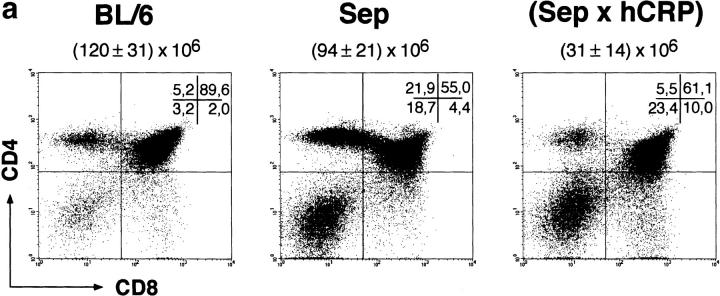
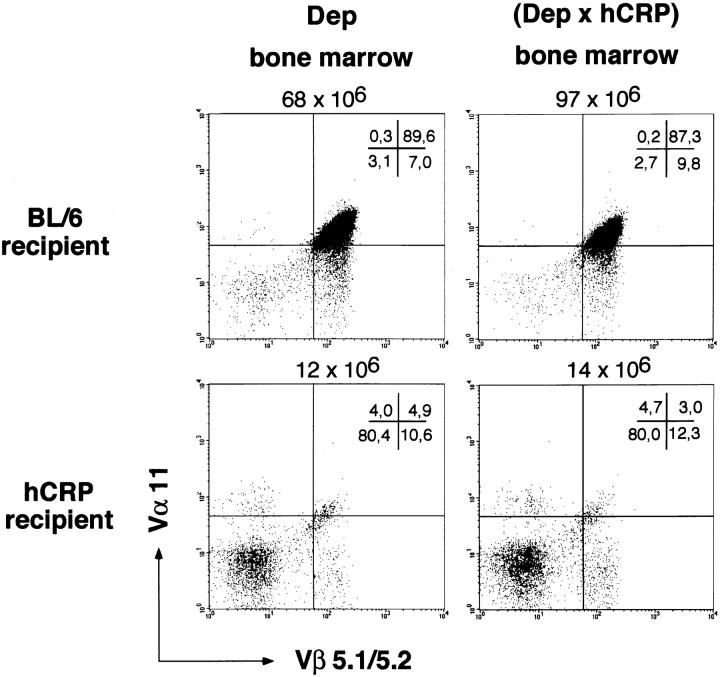
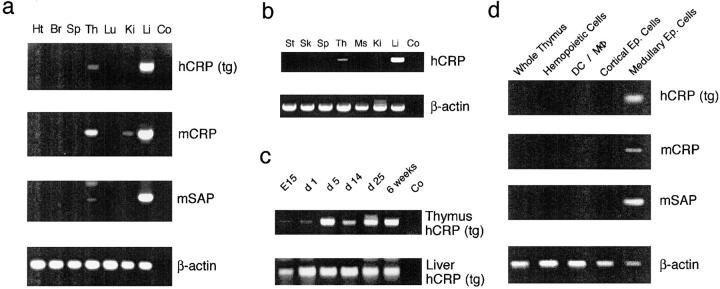
Inducible serum proteins whose concentrations oscillate between nontolerogenic and tolerogenic levels pose a particular challenge to the maintenance of self-tolerance. Temporal restrictions of intrathymic antigen supply should prevent continuous central tolerization of T cells, in analogy to the spatial limitation imposed by tissue-restricted antigen expression. Major acute-phase proteins such as human C-reactive protein (hCRP) are typical examples for such inducible self-antigens. The circulating concentration of hCRP, which is secreted by hepatocytes, is induced up to 1,000-fold during an acute-phase reaction. We have analyzed tolerance to hCRP expressed in transgenic mice under its autologous regulatory regions. Physiological regulation of basal levels (<10(-9) M) and inducibility (>500-fold) are preserved in female transgenics, whereas male transgenics constitutively display induced levels. Surprisingly, crossing of hCRP transgenic mice to two lines of T cell receptor transgenic mice (specific for either a dominant or a subdominant epitope) showed that tolerance is mediated by intrathymic deletion of immature thymocytes, irrespective of widely differing serum levels. In the absence of induction, hCRP expressed by thymic medullary epithelial cells rather than liver-derived hCRP is necessary and sufficient to induce tolerance. Importantly, medullary epithelial cells also express two homologous mouse acute-phase proteins. These results support a physiological role of "ectopic" thymic expression in tolerance induction to acute-phase proteins and possibly other inducible self-antigens and have implications for delineating the relative contributions of central versus peripheral tolerance.
Figures







References
-
- Kappler JW, Roehm N, Marrack P. T cell tolerance by clonal elimination in the thymus. Cell. 1987;49:273–280. - PubMed
-
- Kyewski BA, Fathman CG, Kaplan HS. Intrathymic presentation of circulating non–major histocompatibility complex antigens. Nature. 1984;308:196–199. - PubMed
-
- Arnold, B., G. Schönrich, I. Ferber, J. Alferink, and G.J. Hämmerling. 1994. Peripheral T cell tolerance: distinct levels and multistep mechanisms. In Transgenesis and Targeted Mutagenesis in Immunology. H. Bluethmann and P.S. Ohashi, editors. Academic Press, San Diego, CA. 135–140.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
