

The coordinated action of CC chemokines in the lung orchestrates allergic inflammation and airway hyperresponsiveness
- PMID: 9653092
- PMCID: PMC2525544
- DOI: 10.1084/jem.188.1.157
The coordinated action of CC chemokines in the lung orchestrates allergic inflammation and airway hyperresponsiveness
Abstract
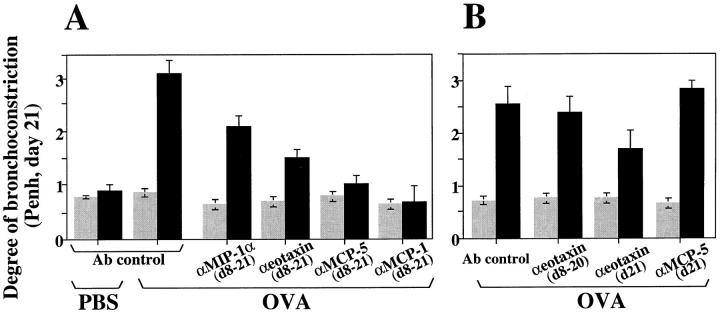
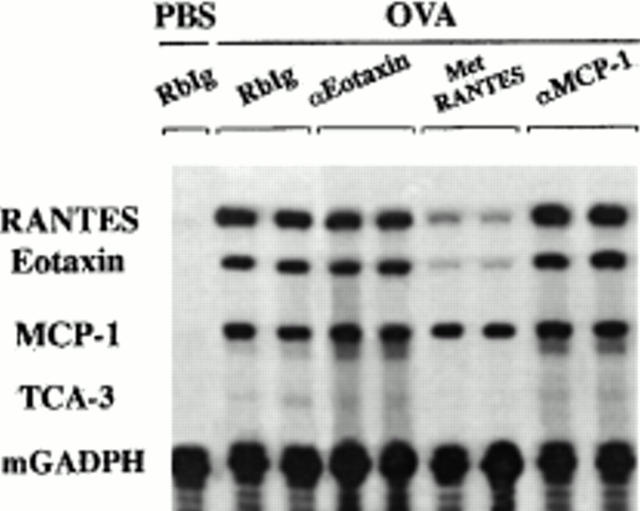
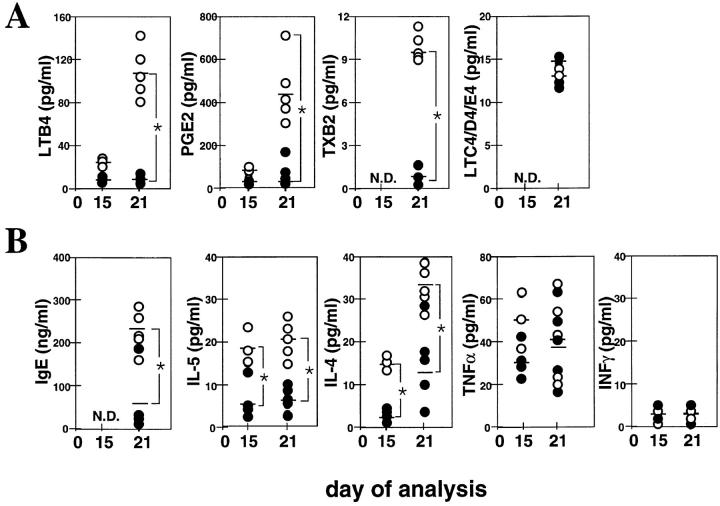
The complex pathophysiology of lung allergic inflammation and bronchial hyperresponsiveness (BHR) that characterize asthma is achieved by the regulated accumulation and activation of different leukocyte subsets in the lung. The development and maintenance of these processes correlate with the coordinated production of chemokines. Here, we have assessed the role that different chemokines play in lung allergic inflammation and BHR by blocking their activities in vivo. Our results show that blockage of each one of these chemokines reduces both lung leukocyte infiltration and BHR in a substantially different way. Thus, eotaxin neutralization reduces specifically BHR and lung eosinophilia transiently after each antigen exposure. Monocyte chemoattractant protein (MCP)-5 neutralization abolishes BHR not by affecting the accumulation of inflammatory leukocytes in the airways, but rather by altering the trafficking of the eosinophils and other leukocytes through the lung interstitium. Neutralization of RANTES (regulated upon activation, normal T cell expressed and secreted) receptor(s) with a receptor antagonist decreases significantly lymphocyte and eosinophil infiltration as well as mRNA expression of eotaxin and RANTES. In contrast, neutralization of one of the ligands for RANTES receptors, macrophage-inflammatory protein 1alpha, reduces only slightly lung eosinophilia and BHR. Finally, MCP-1 neutralization diminishes drastically BHR and inflammation, and this correlates with a pronounced decrease in monocyte- and lymphocyte-derived inflammatory mediators. These results suggest that different chemokines activate different cellular and molecular pathways that in a coordinated fashion contribute to the complex pathophysiology of asthma, and that their individual blockage results in intervention at different levels of these processes.
Figures

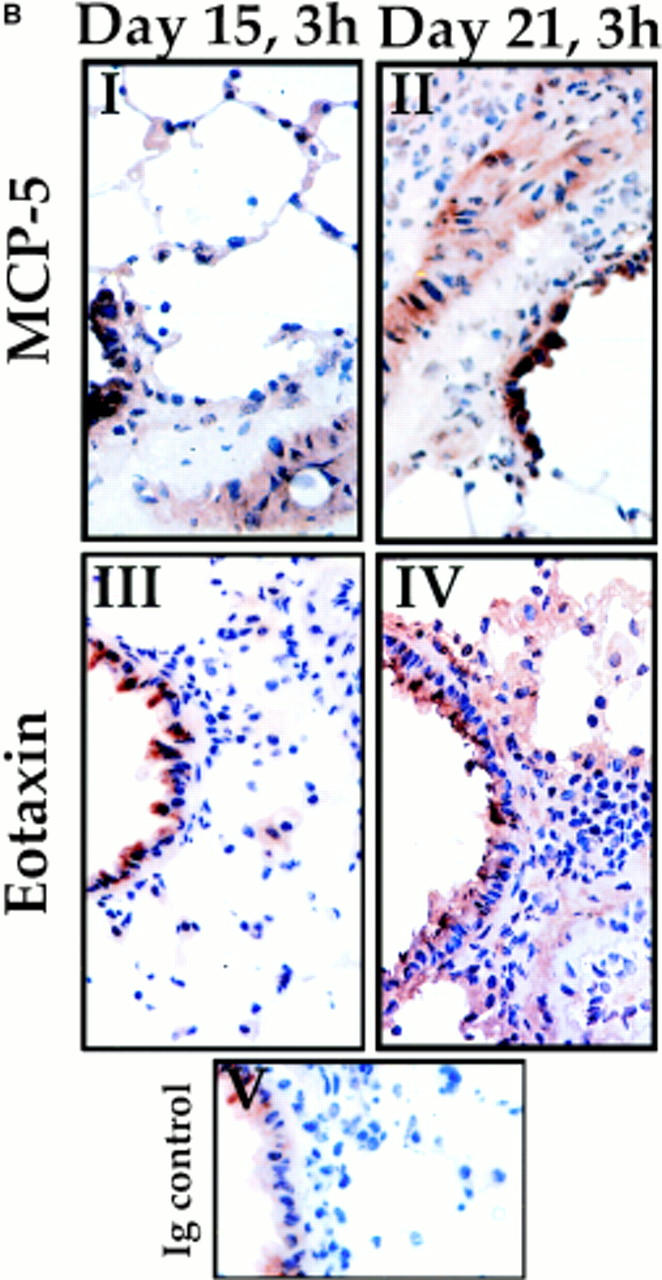
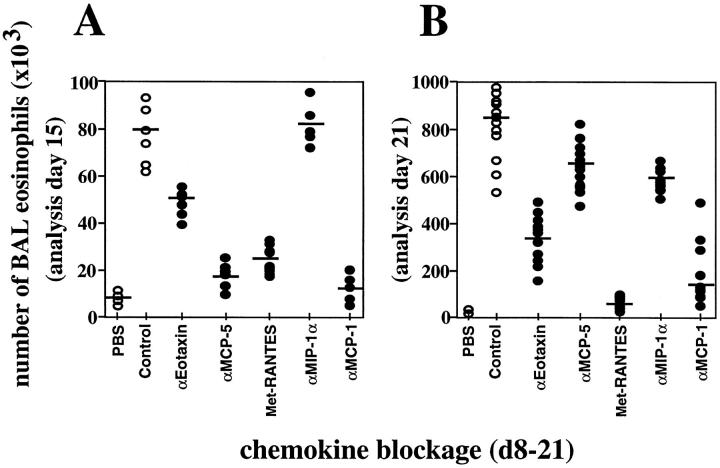
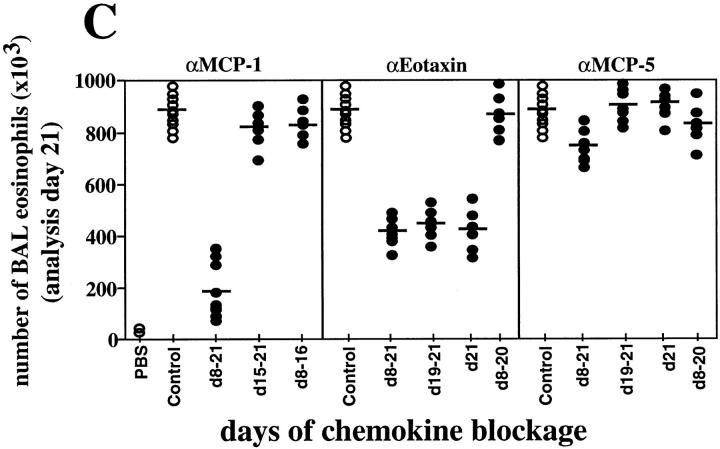
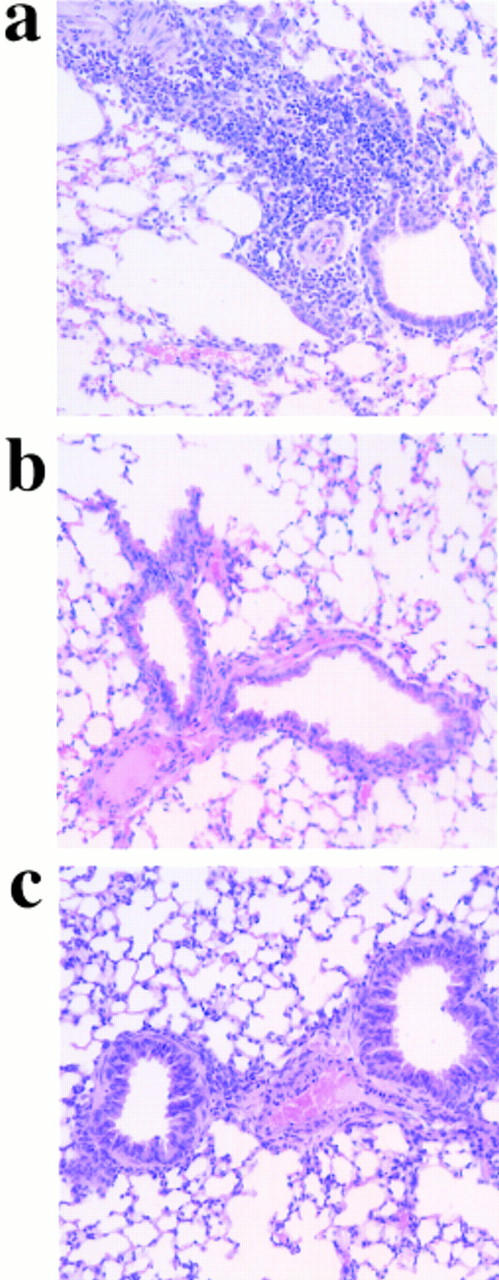
References
-
- Gleich GJ, Flavahan NA, Fujisawa T, Vanhoutte PM. The eosinophil as a mediator of damage to respiratory epithelium: a model for bronchial hyperreactivity. J Allergy Clin Immunol. 1988;81:776–781. - PubMed
-
- Gleich GJ. The eosinophil and bronchial asthma. Current understanding. J Allergy Clin Immunol. 1990;85:422–436. - PubMed
-
- Bradley BL, Azzawi M, Jacobson M, Assoufi B, Collins JV, Irani A-MA, Schwartz LB, Durham SR, Jeffrey PK, Kay AB. Eosinophils, T-lymphocytes, mast cells, neutrophils and macrophages in bronchial biopsy specimens from atopic subjects with asthma: comparison with biopsy specimens from atopic subjects without asthma and normal control subjects and relationship to bronchial hyperresponsiveness. J Allergy Clin Immunol. 1991;88:661–667. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
