

Ilarviruses encode a Cucumovirus-like 2b gene that is absent in other genera within the Bromoviridae
- PMID: 9658153
- PMCID: PMC109913
- DOI: 10.1128/JVI.72.8.6956-6959.1998
Ilarviruses encode a Cucumovirus-like 2b gene that is absent in other genera within the Bromoviridae
Abstract
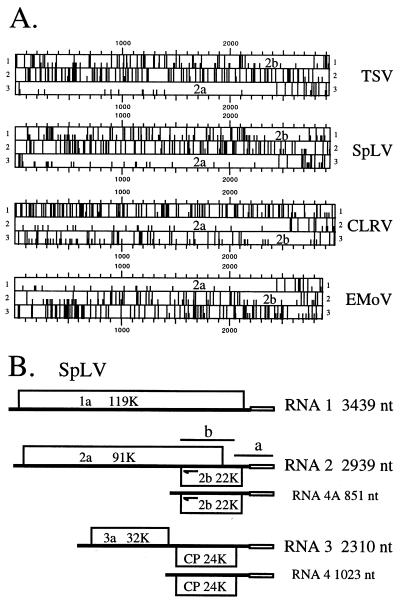
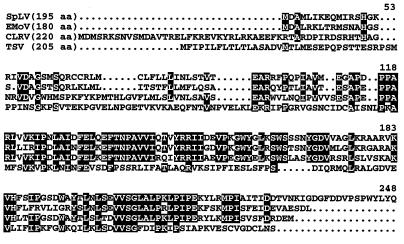
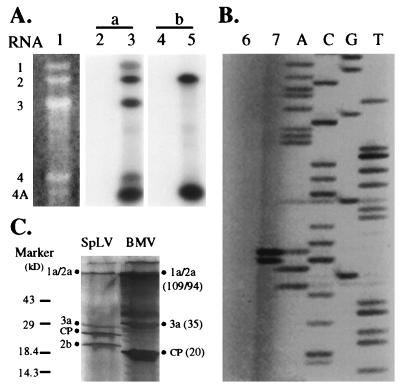
We found that RNA 2 of the four ilarviruses sequenced to date encodes an additional conserved open reading frame (ORF), 2b, that overlaps the 3' end of the previously known ORF, 2a. A novel RNA species of 851 nucleotides was found to accumulate to high levels in plants infected with spinach latent virus (SpLV). Further analysis showed that RNA 4A is a subgenomic RNA of RNA 2 and encodes all of ORF 2b. Moreover, a protein species of the size expected for SpLV ORF 2b was translated in vitro from the RNA 4A-containing virion RNAs. The data support the suggestion that the SpLV 2b protein is translated in vivo. The 2b gene of ilarviruses, which is not encoded by alfamoviruses and bromoviruses, shares several features with the previously reported cucumovirus 2b gene; however, their encoded proteins share no detectable sequence similarities. The evolutionary origin of the 2b gene is discussed.
Figures
References
-
- Bachman E J, Scott S W, Xin G, Vance V B. The complete nucleotide sequence of prune dwarf ilarvirus RNA 3: implications for coat protein activation of genome replication in ilarviruses. Virology. 1994;201:127–131. - PubMed
-
- Bol J F, van Vloten-Doting L, Jaspars E M. A functional equivalence of top component a RNA and coat protein in the initiation of infection by alfalfa mosaic virus. Virology. 1971;46:73–85. - PubMed
-
- Bos L. Spinach latent virus. CMI/ABB descriptions of plant viruses, no. 281. Warwick, United Kingdom: Association of Applied Biologists; 1984.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
