

Isolation and contraction of the stress fiber
- PMID: 9658180
- PMCID: PMC25437
- DOI: 10.1091/mbc.9.7.1919
Isolation and contraction of the stress fiber
Abstract
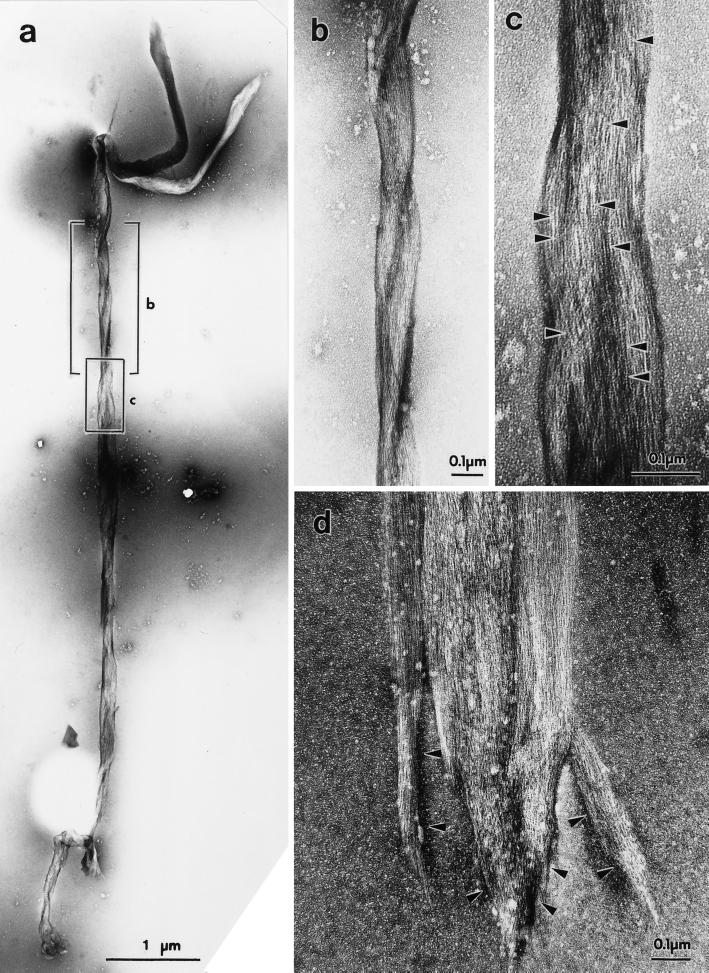
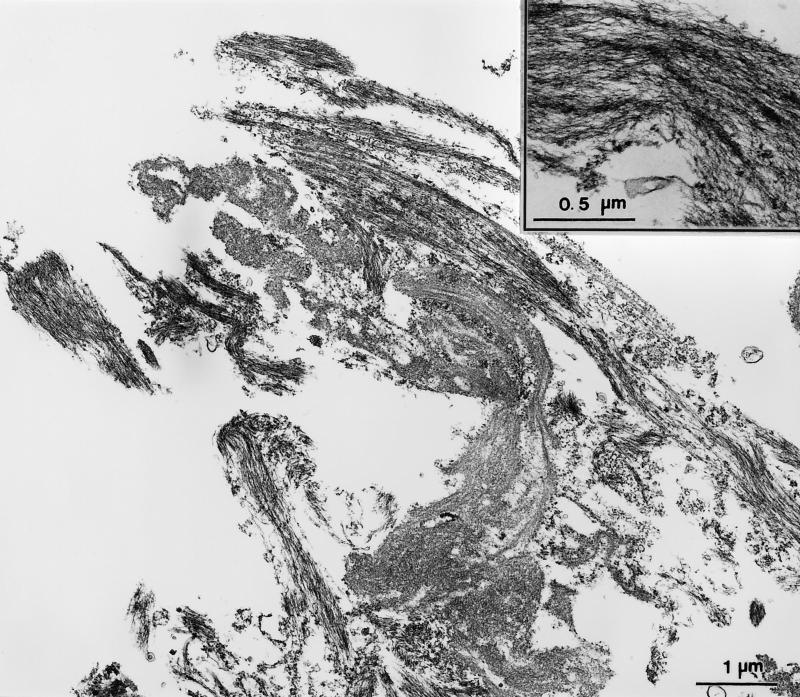
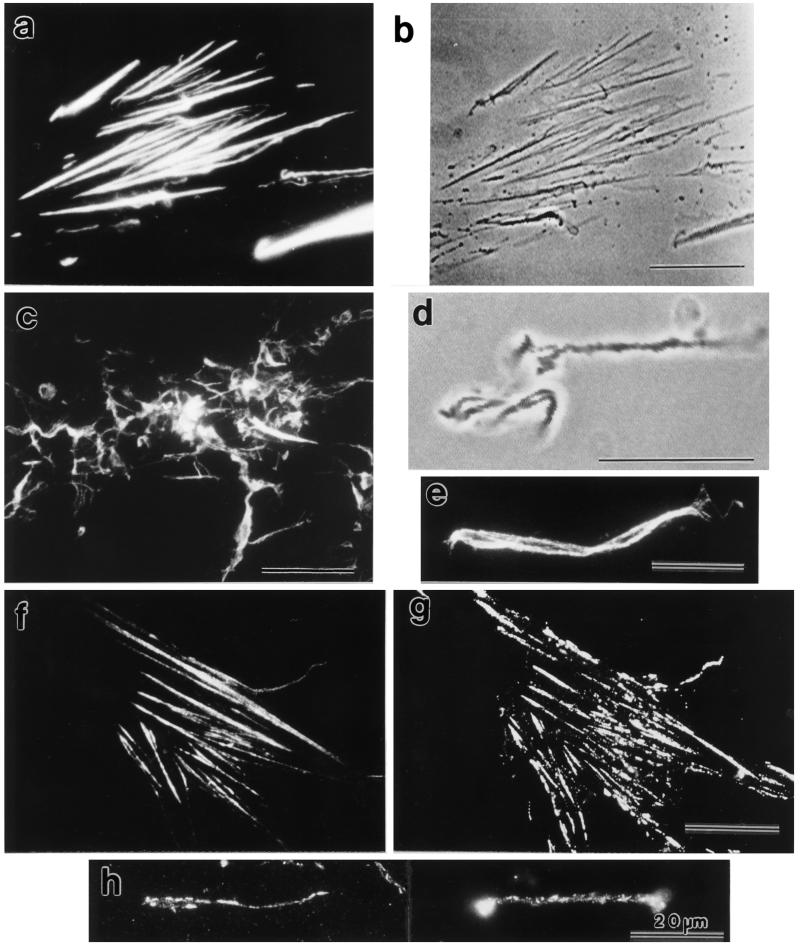
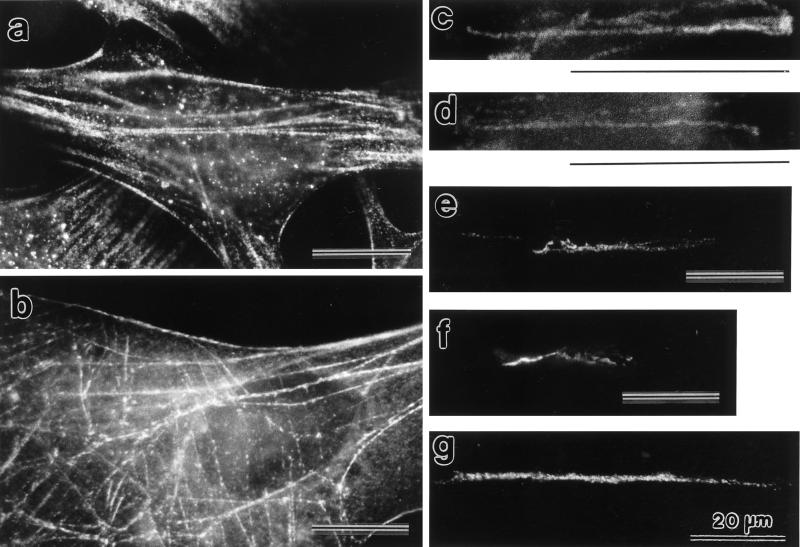
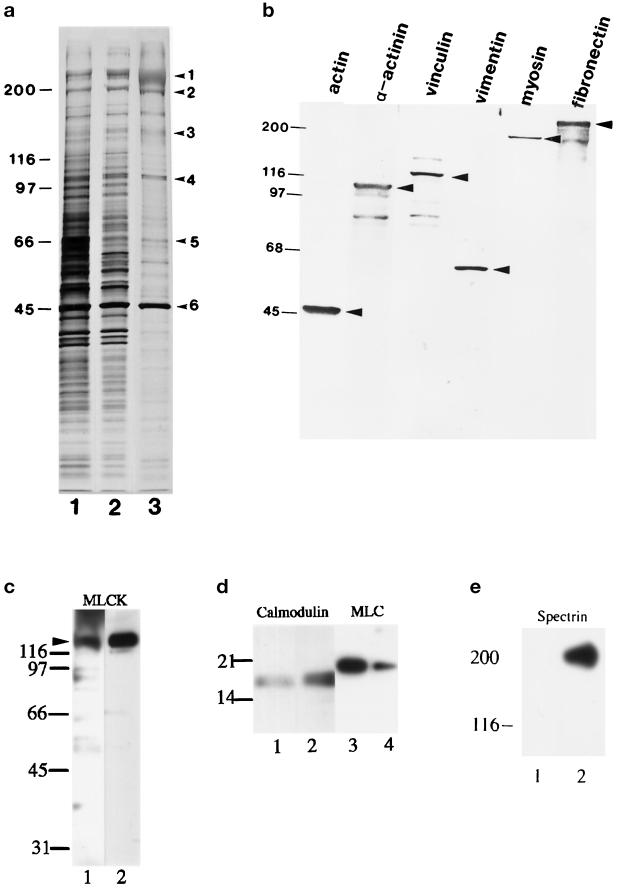
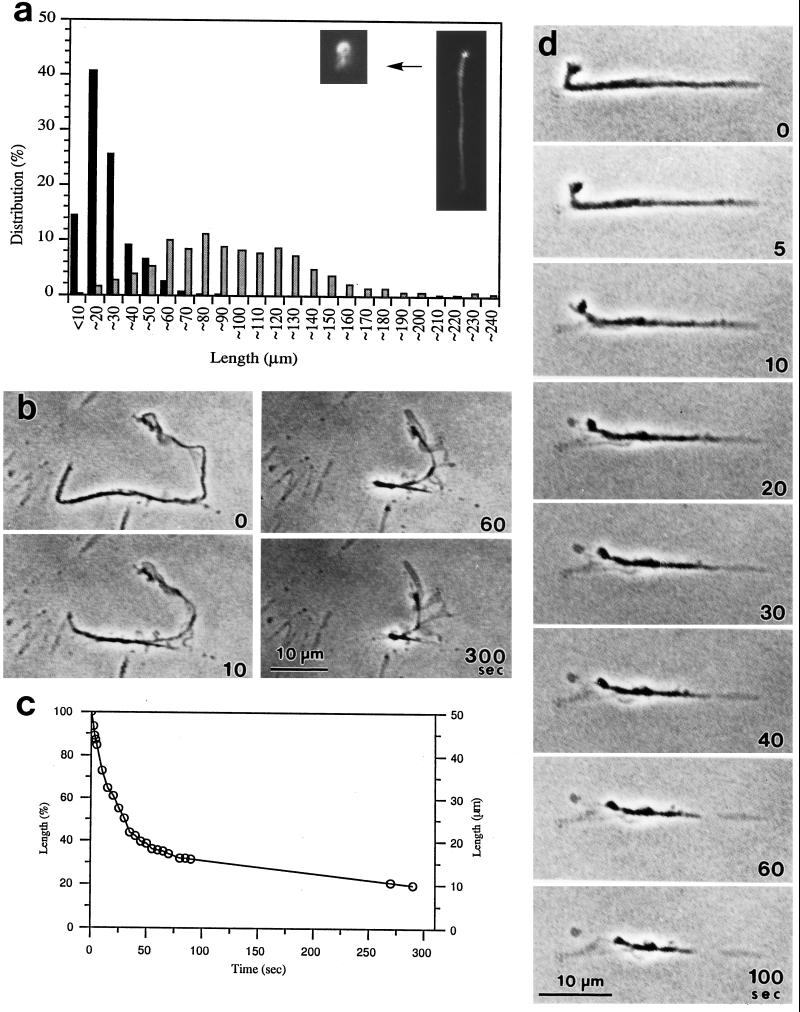
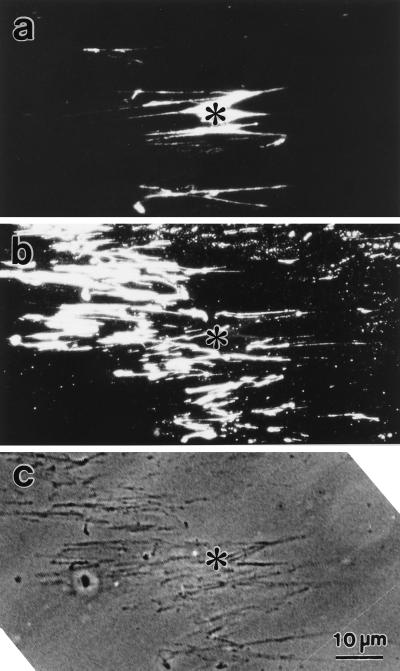
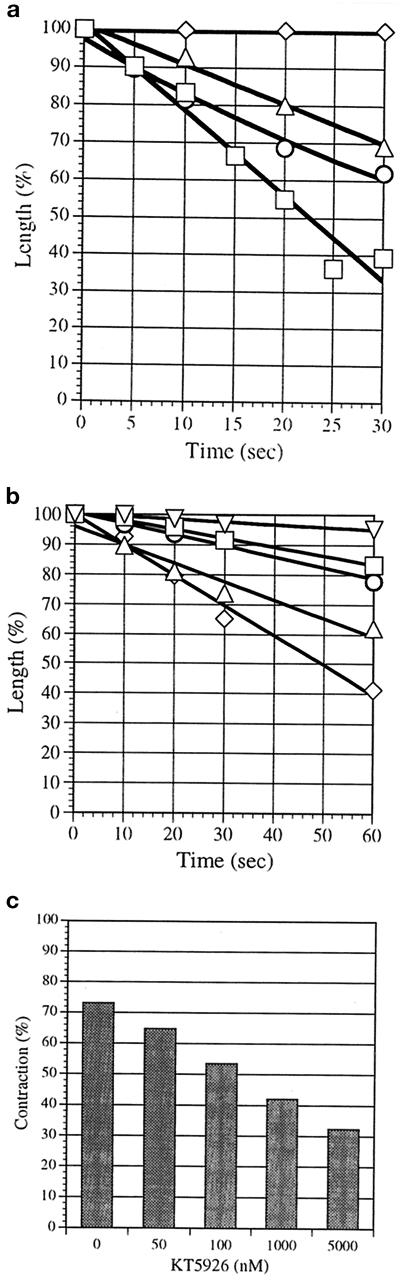
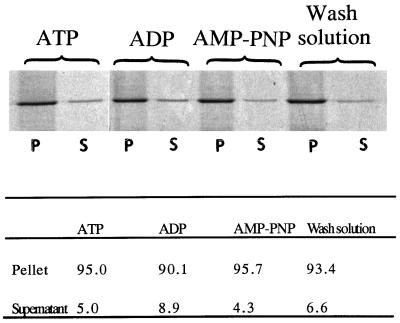
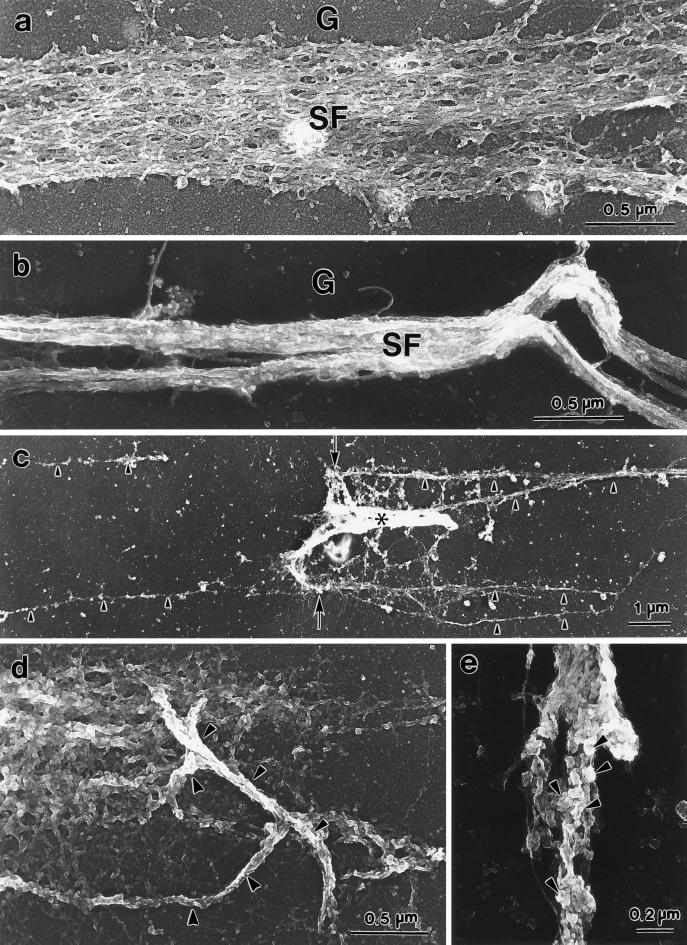
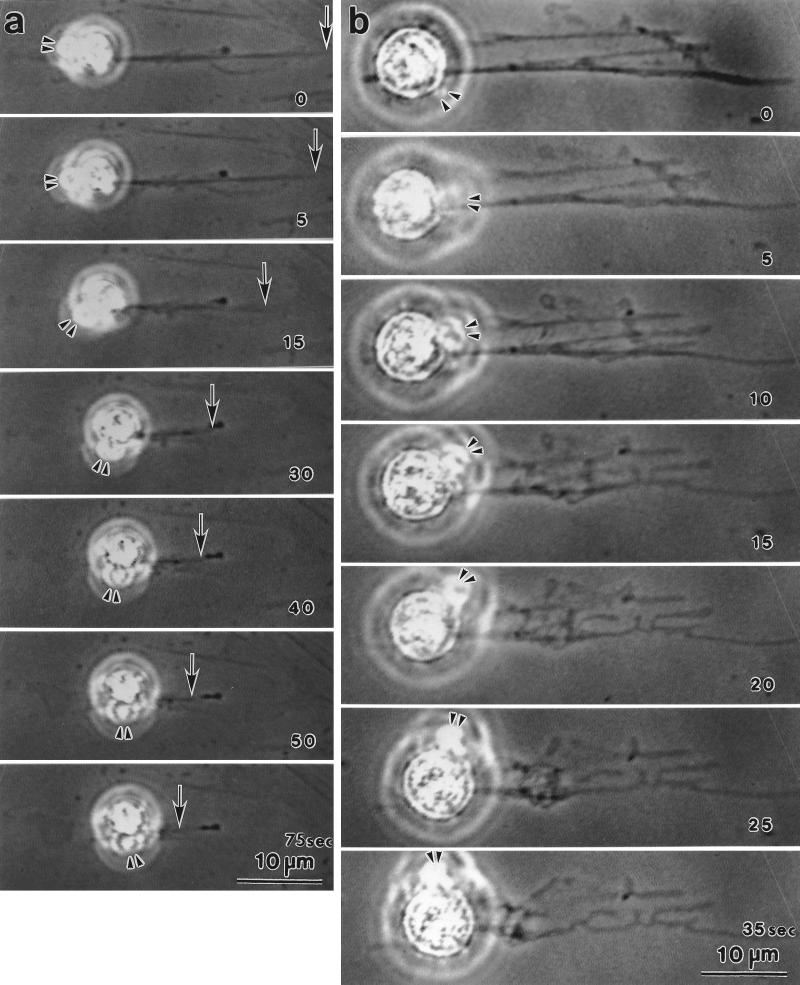
Stress fibers were isolated from cultured human foreskin fibroblasts and bovine endothelial cells, and their contraction was demonstrated in vitro. Cells in culture dishes were first treated with a low-ionic-strength extraction solution and then further extracted using detergents. With gentle washes by pipetting, the nucleus and the apical part of cells were removed. The material on the culture dish was scraped, and the freed material was forced through a hypodermic needle and fractionated by sucrose gradient centrifugation. Isolated, free-floating stress fibers stained brightly with fluorescently labeled phalloidin. When stained with anti-alpha-actinin or anti-myosin, isolated stress fibers showed banded staining patterns. By electron microscopy, they consisted of bundles of microfilaments, and electron-dense areas were associated with them in a semiperiodic manner. By negative staining, isolated stress fibers often exhibited gentle twisting of microfilament bundles. Focal adhesion-associated proteins were also detected in the isolated stress fiber by both immunocytochemical and biochemical means. In the presence of Mg-ATP, isolated stress fibers shortened, on the average, to 23% of the initial length. The maximum velocity of shortening was several micrometers per second. Polystyrene beads on shortening isolated stress fibers rotated, indicating spiral contraction of stress fibers. Myosin regulatory light chain phosphorylation was detected in contracting stress fibers, and a myosin light chain kinase inhibitor, KT5926, inhibited isolated stress fiber contraction. Our study demonstrates that stress fibers can be isolated with no apparent loss of morphological features and that they are truly contractile organelle.
Figures

References
-
- Avnur Z, Geiger B. Substrate attached membranes of cultured cells. Isolation and characterization of ventral membranes and the associated cytoskeleton. J Mol Biol. 1981;153:361–379. - PubMed
-
- Badley RA, Lloyd CW, Woods A, Carruthers L, Allcock C. Mechanisms of cellular adhesion. Exp Cell Res. 1978;117:231–244. - PubMed
-
- Brands R, Feltkamp CA. Wet cleaving of cell: a method to introduce macromolecules into the cytoplasm. Exp Cell Res. 1988;176:309–318. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
