

The human homologue of Bub3 is required for kinetochore localization of Bub1 and a Mad3/Bub1-related protein kinase
- PMID: 9660858
- PMCID: PMC2133037
- DOI: 10.1083/jcb.142.1.1
The human homologue of Bub3 is required for kinetochore localization of Bub1 and a Mad3/Bub1-related protein kinase
Abstract
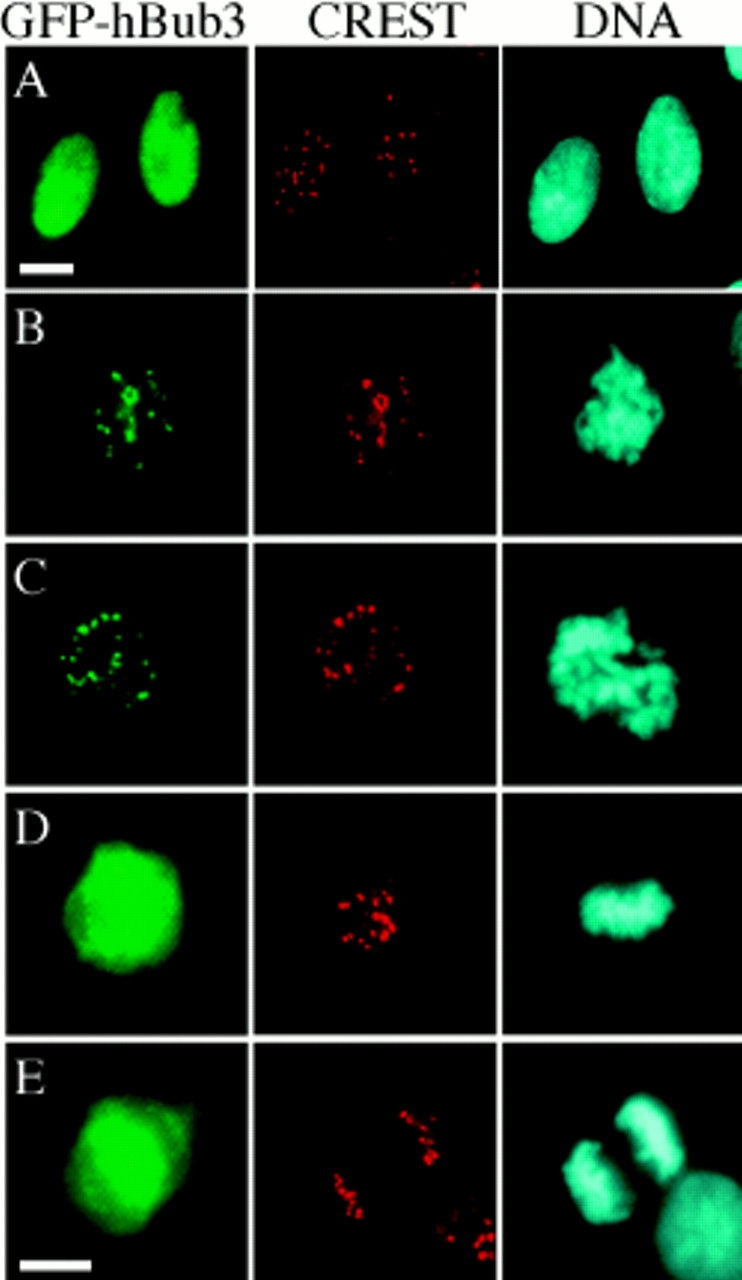
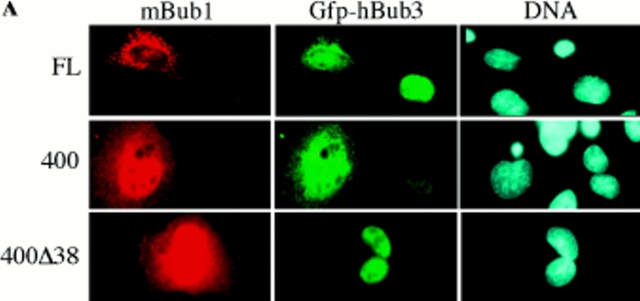
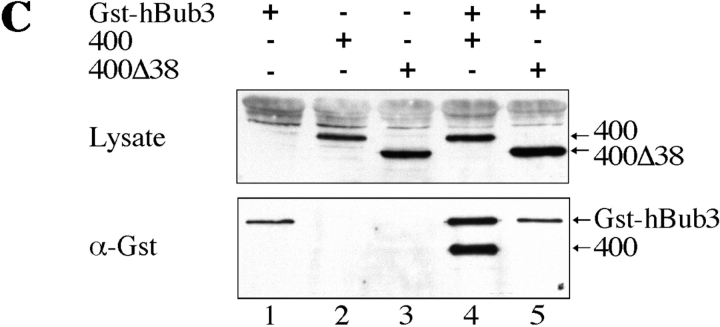
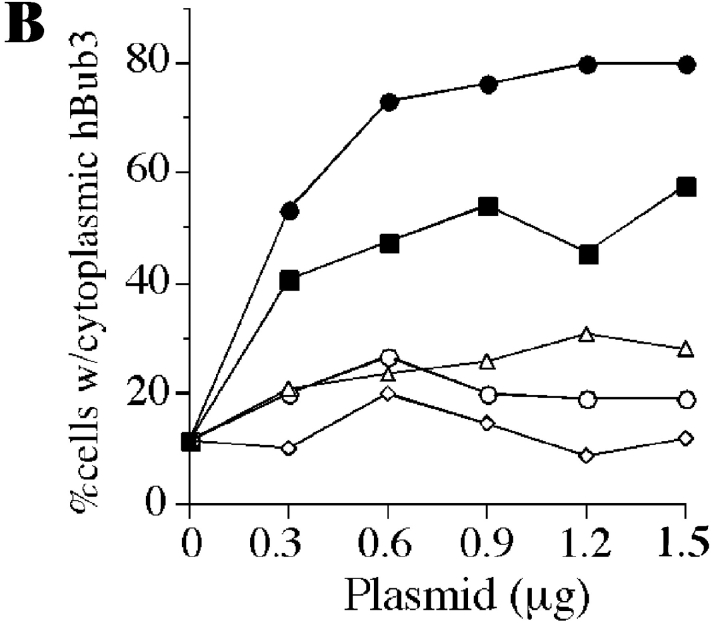
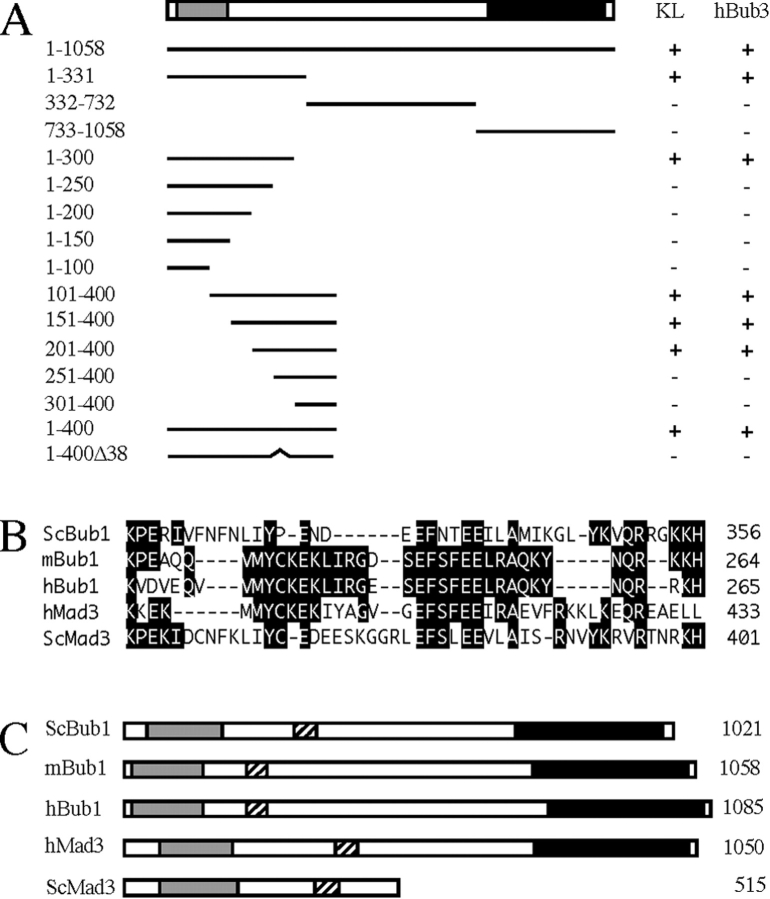
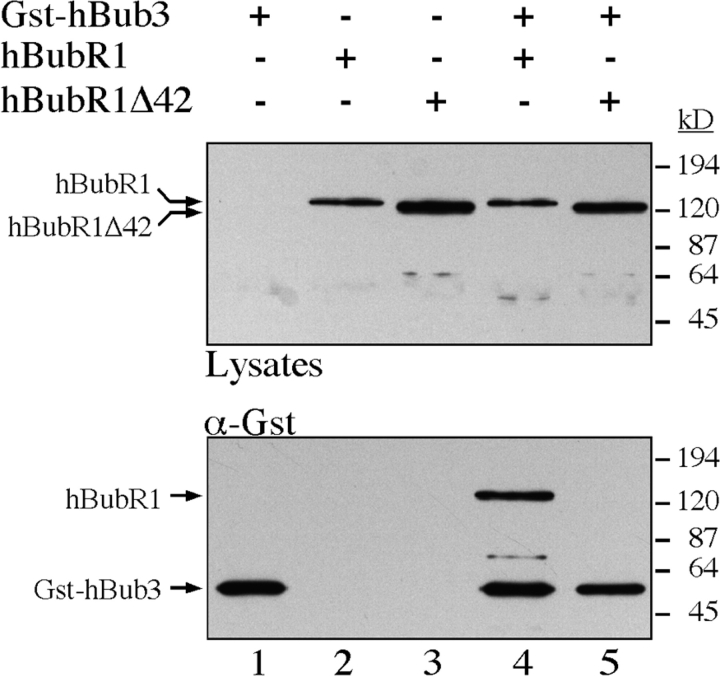
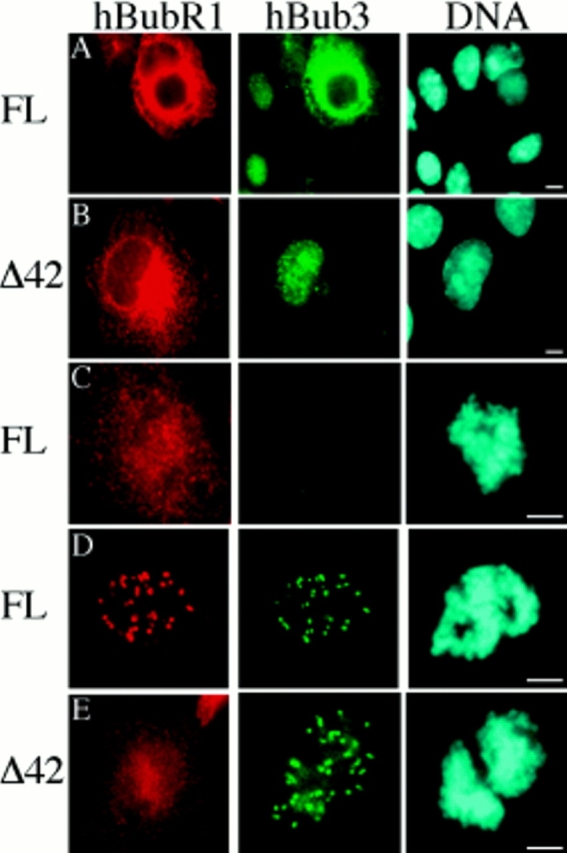
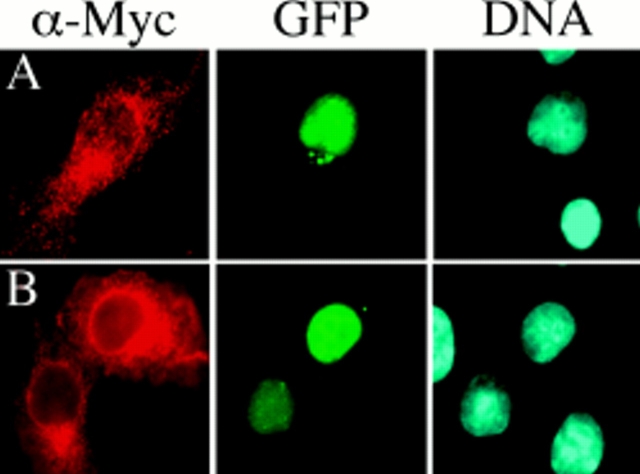
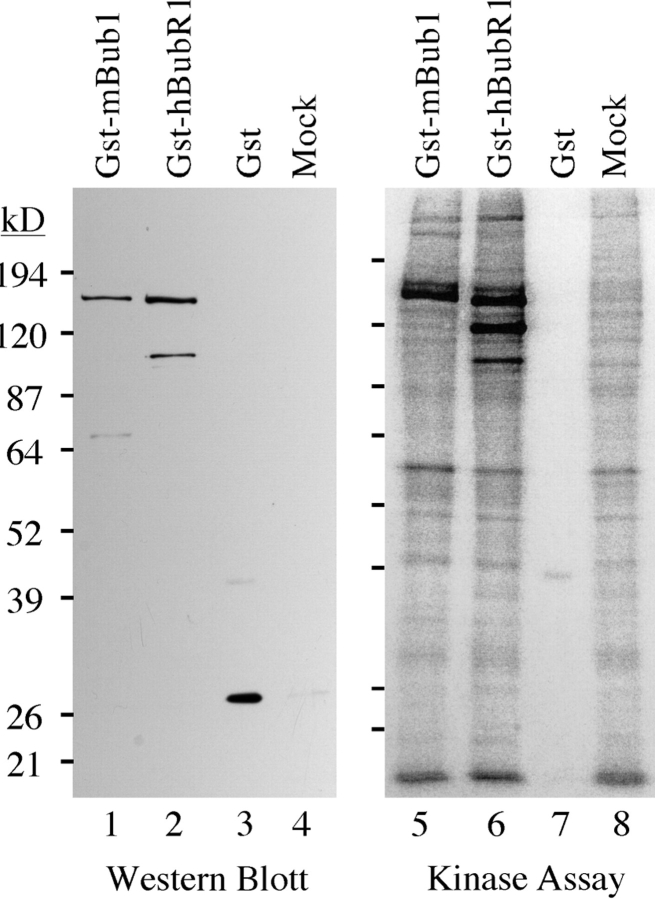
A feedback control mechanism, or cell cycle checkpoint, delays the onset of anaphase until all the chromosomes are correctly aligned on the mitotic spindle. Previously, we showed that the murine homologue of Bub1 is not only required for checkpoint response to spindle damage, but also restrains progression through a normal mitosis (Taylor, S.S., and F. McKeon. 1997. Cell. 89:727-735). Here, we describe the identification of a human homologue of Bub3, a 37-kD protein with four WD repeats. Like Bub1, Bub3 localizes to kinetochores before chromosome alignment. In addition, Bub3 and Bub1 interact in mammalian cells. Deletion mapping was used to identify the domain of Bub1 required for binding Bub3. Significantly, this same domain is required for kinetochore localization of Bub1, suggesting that the role of Bub3 is to localize Bub1 to the kinetochore, thereby activating the checkpoint in response to unattached kinetochores. The identification of a human Mad3/Bub1-related protein kinase, hBubR1, which can also bind Bub3 in mammalian cells, is described. Ectopically expressed hBubR1 also localizes to kinetochores during prometaphase, but only when hBub3 is overexpressed. We discuss the implications of the common interaction between Bub1 and hBubR1 with hBub3 for checkpoint control.
Figures


References
-
- Adams MD, Kerlavage AR, Fleischmann RD, Fuldner RA, Bult CJ, Lee NH, Kirkness EF, Weinstock KG, Gocayne JD, White O, et al. Initial assessment of human gene diversity and expression patterns based upon 83 million nucleotides of cDNA sequence. Nature. 1995;377(Suppl.):3–174. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Bharathi A, Ghosh A, Whalen WA, Yoon JH, Pu R, Dasso M, Dhar R. The human RAE1 gene is a functional homologue of Schizosaccharomyces pombe rae1gene involved in nuclear export of Poly(A)+ RNA. Gene. 1997;198:251–258. - PubMed
-
- Brown JA, Bharathi A, Ghosh A, Whalen W, Fitzgerald E, Dhar R. A mutation in the Schizosaccharomyces pombe rae1gene causes defects in poly(A)+ RNA export and in the cytoskeleton. J Biol Chem. 1995;270:7411–7419. - PubMed
-
- Cahill DP, Lengauer C, Yu J, Riggins GJ, Willson JK, Markowitz SD, Kinzler KW, Vogelstein B. Mutations of mitotic checkpoint genes in human cancers. Nature. 1998;392:300–303. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
