

Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration
- PMID: 9660867
- PMCID: PMC2133024
- DOI: 10.1083/jcb.142.1.117
Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration
Abstract
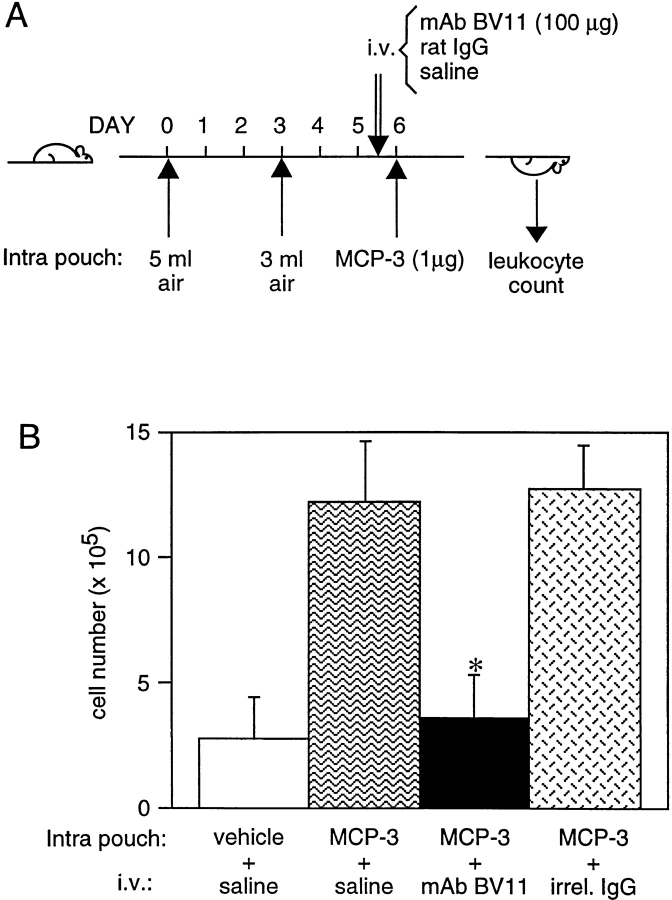
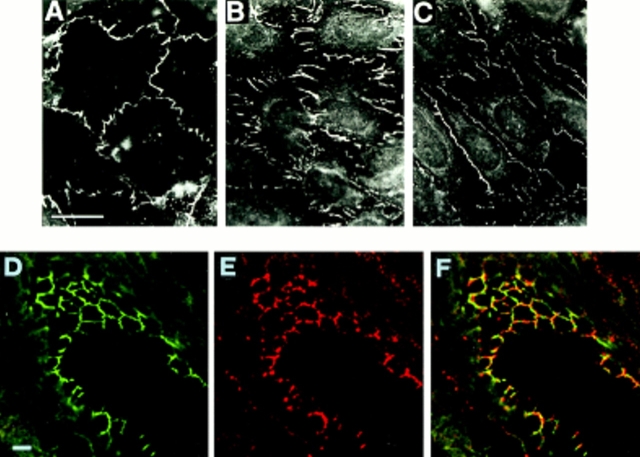
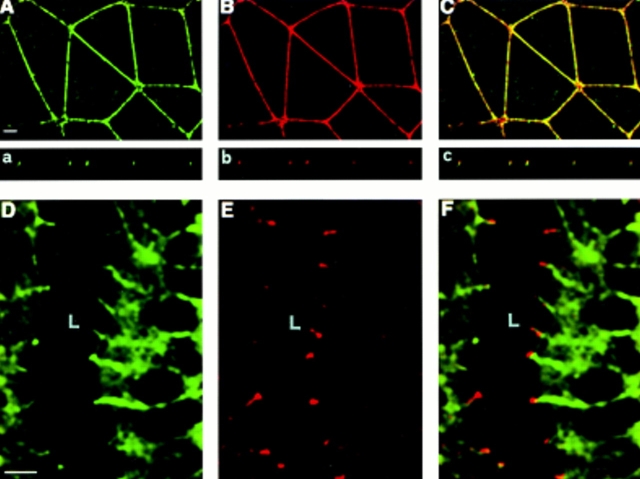
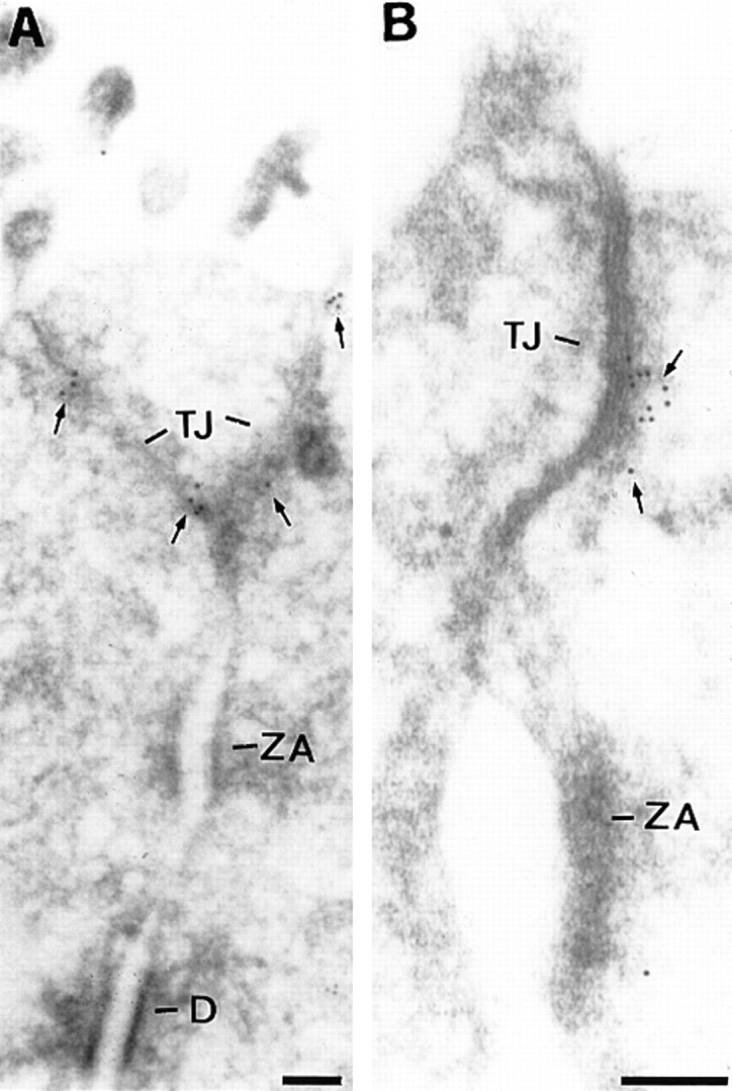
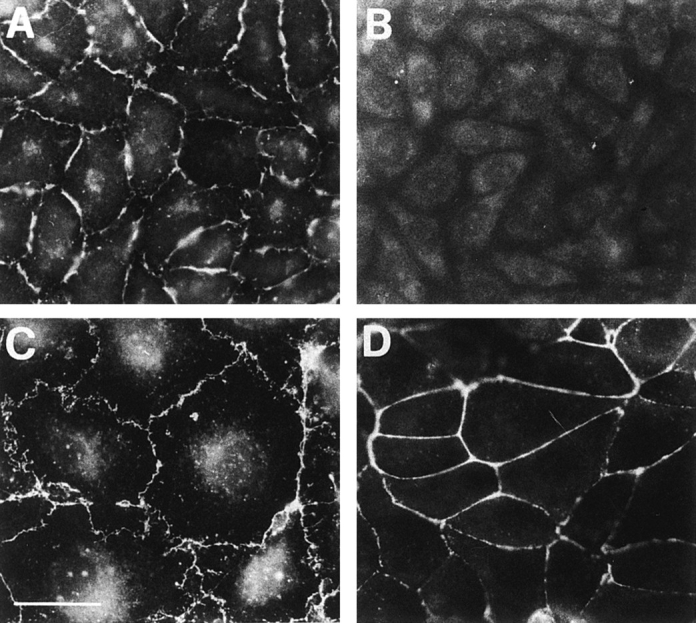
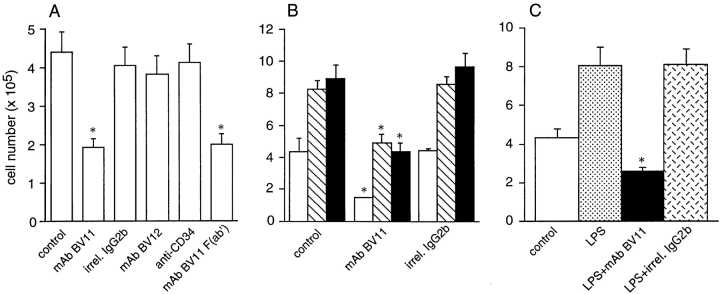
Tight junctions are the most apical components of endothelial and epithelial intercellular cleft. In the endothelium these structures play an important role in the control of paracellular permeability to circulating cells and solutes. The only known integral membrane protein localized at sites of membrane-membrane interaction of tight junctions is occludin, which is linked inside the cells to a complex network of cytoskeletal and signaling proteins. We report here the identification of a novel protein (junctional adhesion molecule [JAM]) that is selectively concentrated at intercellular junctions of endothelial and epithelial cells of different origins. Confocal and immunoelectron microscopy shows that JAM codistributes with tight junction components at the apical region of the intercellular cleft. A cDNA clone encoding JAM defines a novel immunoglobulin gene superfamily member that consists of two V-type Ig domains. An mAb directed to JAM (BV11) was found to inhibit spontaneous and chemokine-induced monocyte transmigration through an endothelial cell monolayer in vitro. Systemic treatment of mice with BV11 mAb blocked monocyte infiltration upon chemokine administration in subcutaneous air pouches. Thus, JAM is a new component of endothelial and epithelial junctions that play a role in regulating monocyte transmigration.
Figures
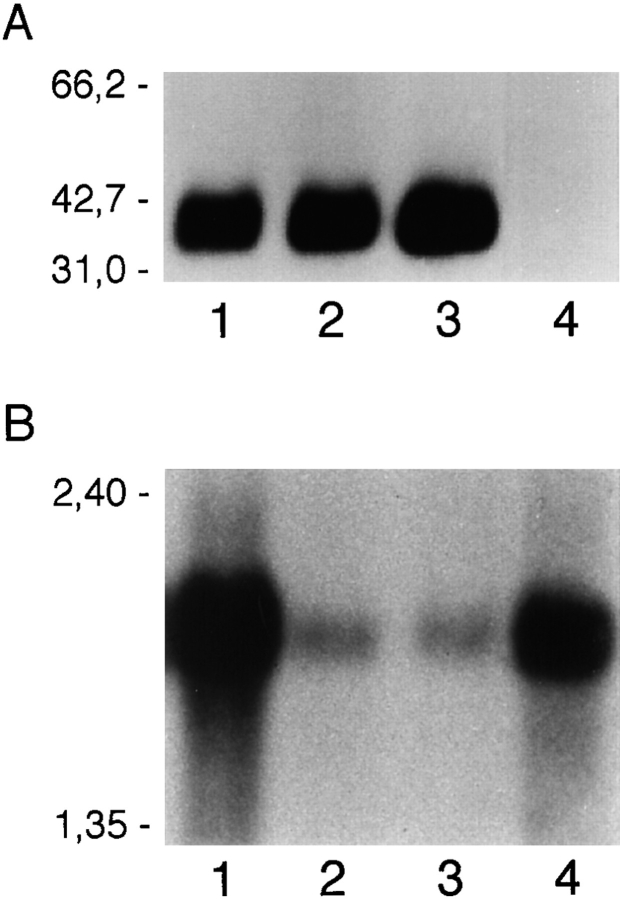
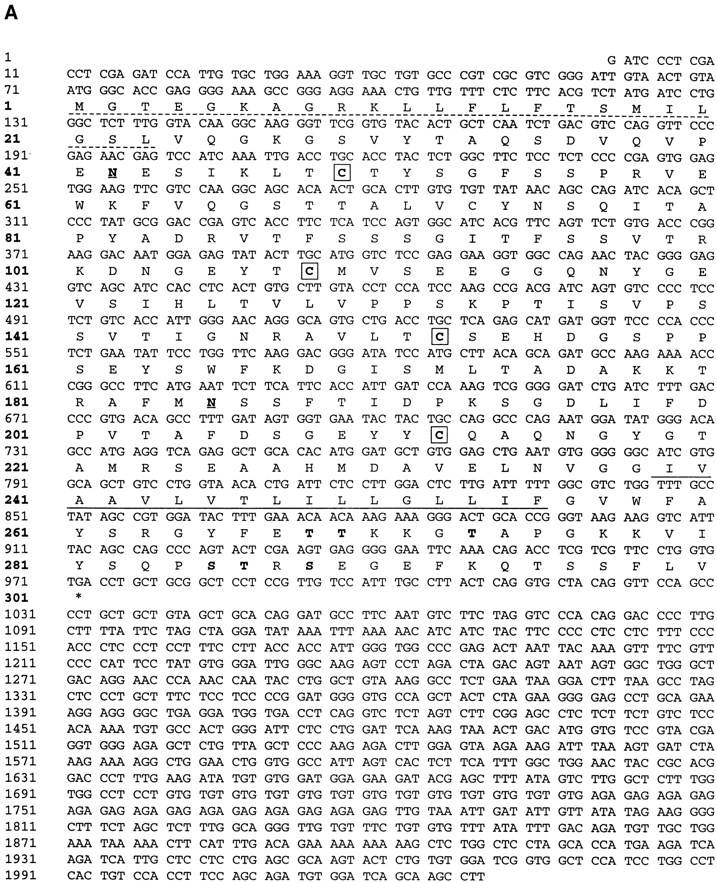
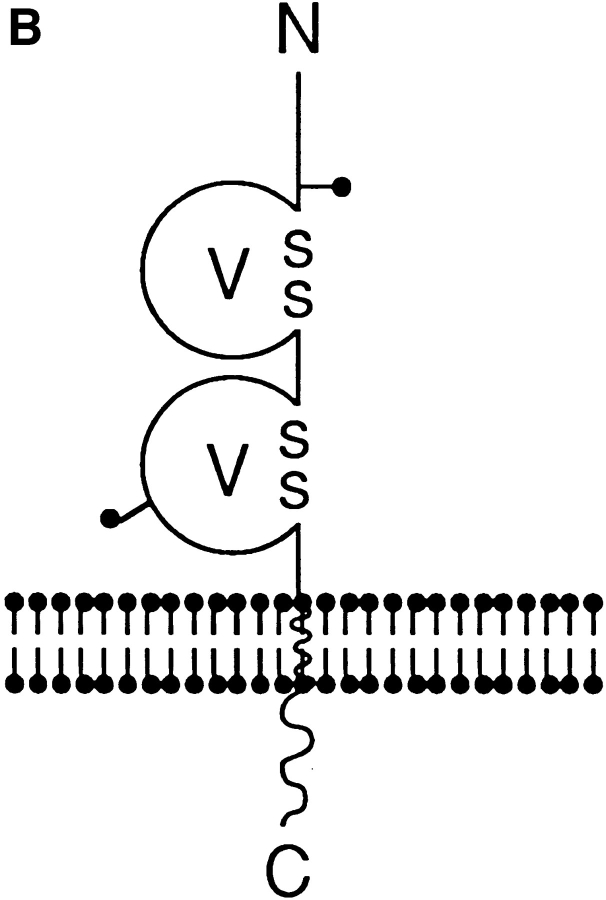
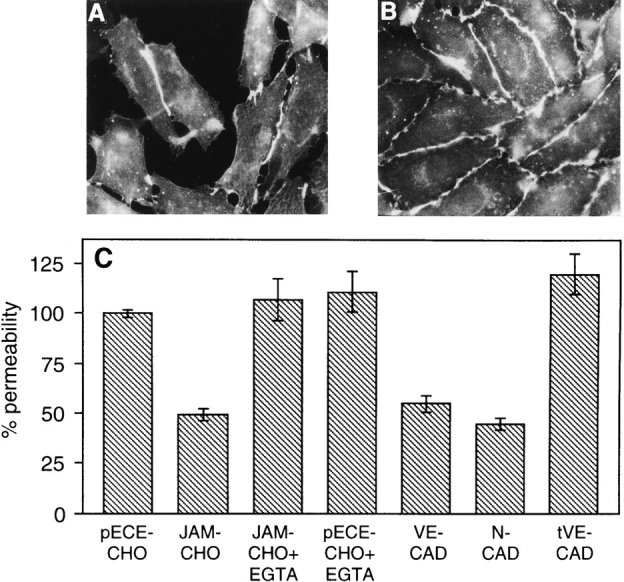
References
-
- Anderson AO, Shaw S. T cell adhesion to endothelium: the FRC conduit system and other anatomic and molecular features which facilitate the adhesion cascade in lymph node. Semin Immunol. 1993;5:271–282. - PubMed
-
- Anderson JM, Balda MS, Fanning AS. The structure and regulation of tight junctions. Curr Opin Cell Biol. 1993;5:772–776. - PubMed
-
- Anderson JM, Van Itallie CM. Tight junctions and the molecular basis for regulation of paracellular permeability. Am J Physiol. 1995;269:G465–G475. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
