

The expression of light-regulated genes in the high-pigment-1 mutant of tomato
- PMID: 9662522
- PMCID: PMC34934
- DOI: 10.1104/pp.117.3.797
The expression of light-regulated genes in the high-pigment-1 mutant of tomato
Abstract
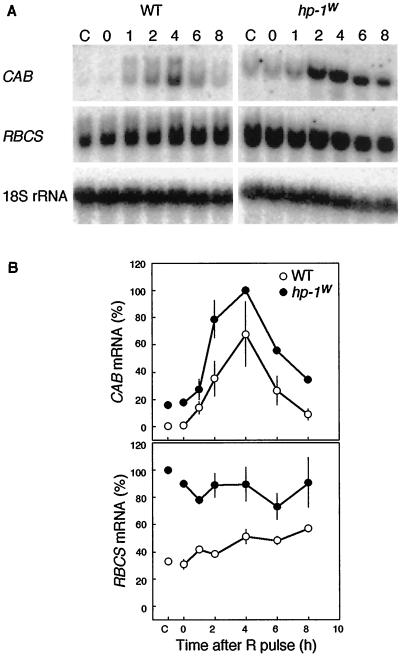
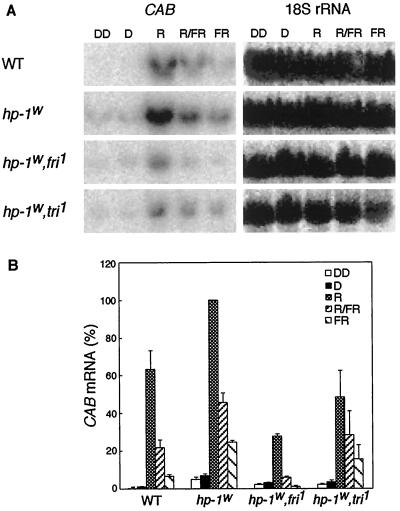
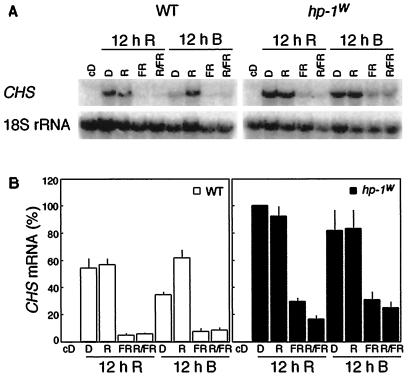
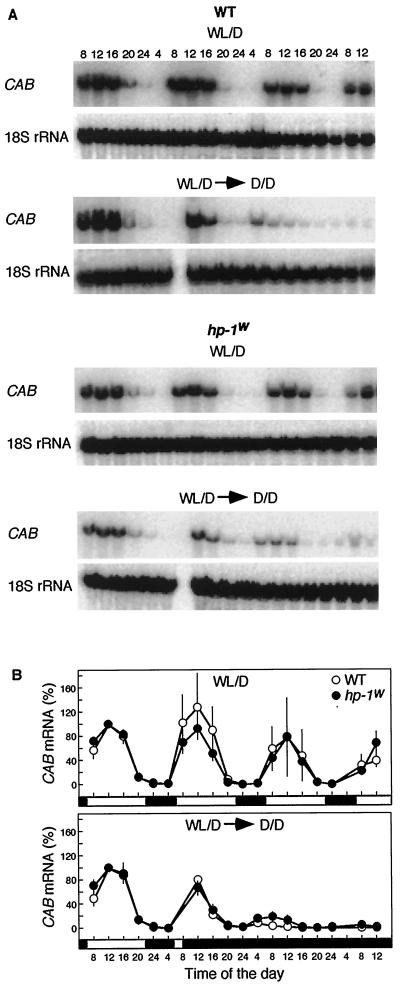
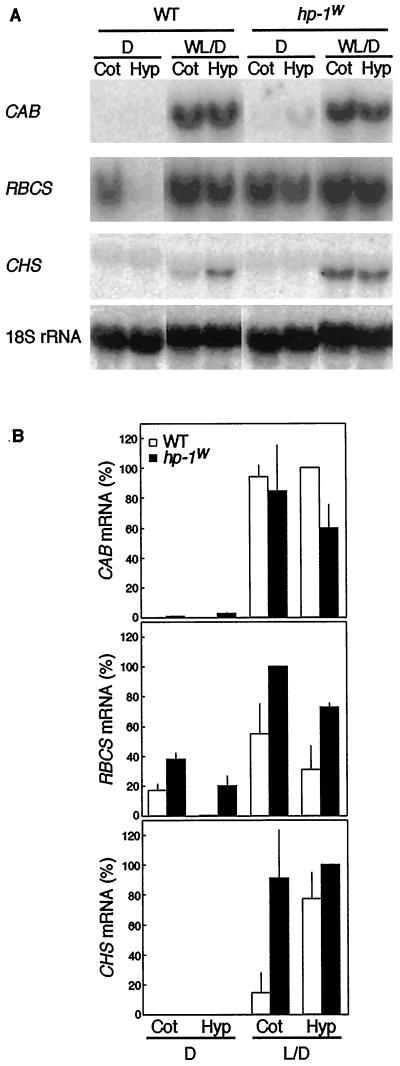
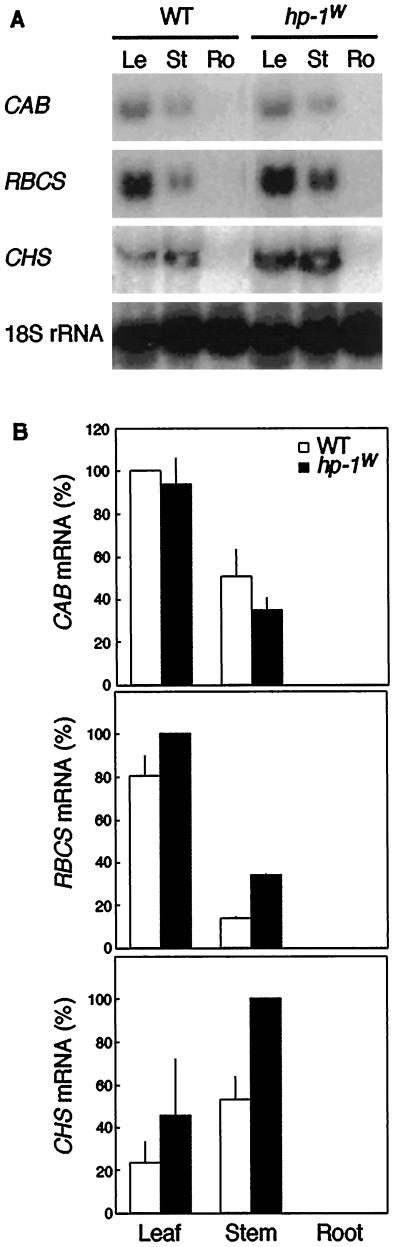
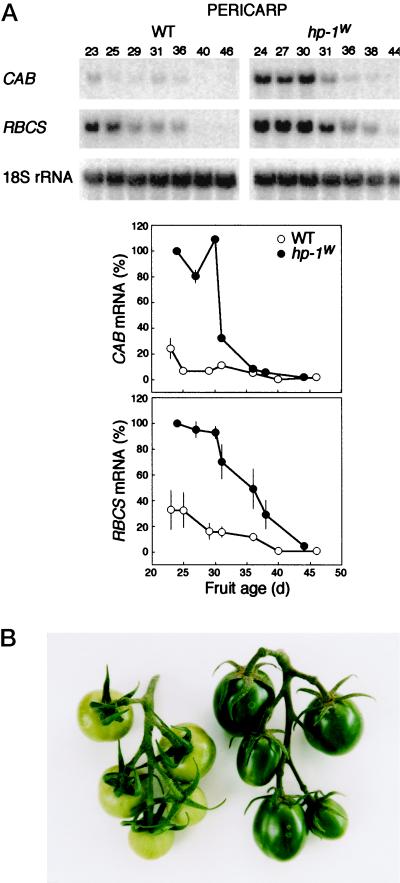
Three light-regulated genes, chlorophyll a/b-binding protein (CAB), ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit, and chalcone synthase (CHS), are demonstrated to be up-regulated in the high-pigment-1 (hp-1) mutant of tomato (Lycopersicon esculentum Mill. ) compared with wild type (WT). However, the pattern of up-regulation of the three genes depends on the light conditions, stage of development, and tissue studied. Compared with WT, the hp-1 mutant showed higher CAB gene expression in the dark after a single red-light pulse and in the pericarp of immature fruits. However, in vegetative tissues of light-grown seedlings and adult plants, CAB mRNA accumulation did not differ between WT and the hp-1 mutant. The ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit mRNA accumulated to a higher level in the hp-1 mutant than WT under all light conditions and tissues studied, whereas CHS gene expression was up-regulated in de-etiolated vegetative hp-1-mutant tissues only. The CAB and CHS genes were shown to be phytochrome regulated and both phytochrome A and B1 play a role in CAB gene expression. These observations support the hypothesis that the HP-1 protein plays a general repressive role in phytochrome signal transduction.
Figures
References
-
- Ahmad M, Cashmore AR. The pef mutants of Arabidopsis thaliana define lesions early in the phytochrome signaling pathway. Plant J. 1996;10:1103–1110. - PubMed
-
- Bowler C, Neuhaus G, Yamagata H, Chua N-H. Cyclic GMP and calcium mediate phytochrome phototransduction. Cell. 1994a;77:73–81. - PubMed
-
- Bowler C, Yamagata H, Neuhaus G, Chua N-H. Phytochrome signal transduction pathways are regulated by reciprocal control mechanisms. Genes Dev. 1994b;8:2188–2202. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
