

Does leaf position within a canopy affect acclimation of photosynthesis to elevated CO2?. Analysis Of a wheat crop under free-air co2 enrichment
- PMID: 9662547
- PMCID: PMC34920
- DOI: 10.1104/pp.117.3.1037
Does leaf position within a canopy affect acclimation of photosynthesis to elevated CO2?. Analysis Of a wheat crop under free-air co2 enrichment
Abstract
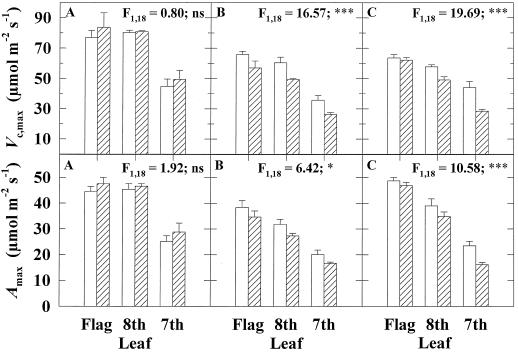
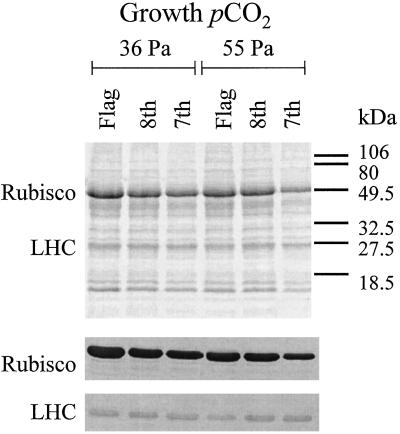
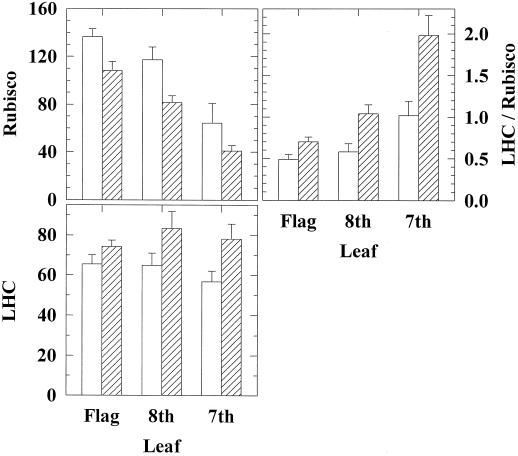
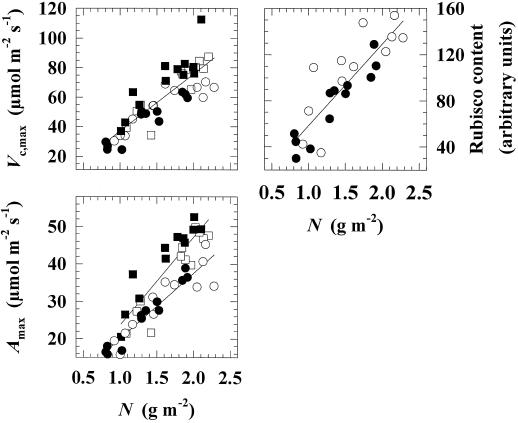
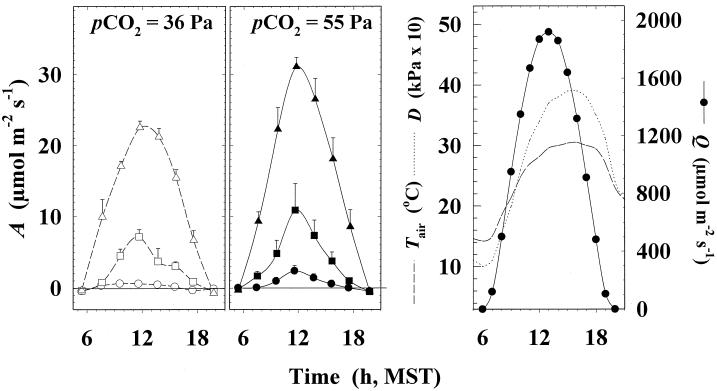
Previous studies of photosynthetic acclimation to elevated CO2 have focused on the most recently expanded, sunlit leaves in the canopy. We examined acclimation in a vertical profile of leaves through a canopy of wheat (Triticum aestivum L.). The crop was grown at an elevated CO2 partial pressure of 55 Pa within a replicated field experiment using free-air CO2 enrichment. Gas exchange was used to estimate in vivo carboxylation capacity and the maximum rate of ribulose-1,5-bisphosphate-limited photosynthesis. Net photosynthetic CO2 uptake was measured for leaves in situ within the canopy. Leaf contents of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), light-harvesting-complex (LHC) proteins, and total N were determined. Elevated CO2 did not affect carboxylation capacity in the most recently expanded leaves but led to a decrease in lower, shaded leaves during grain development. Despite this acclimation, in situ photosynthetic CO2 uptake remained higher under elevated CO2. Acclimation at elevated CO2 was accompanied by decreases in both Rubisco and total leaf N contents and an increase in LHC content. Elevated CO2 led to a larger increase in LHC/Rubisco in lower canopy leaves than in the uppermost leaf. Acclimation of leaf photosynthesis to elevated CO2 therefore depended on both vertical position within the canopy and the developmental stage.
Figures
References
-
- Anderson JM. Photoregulation of the composition, function, and structure of thylakoid membranes. Annu Rev Plant Physiol. 1986;37:93–136.
-
- Baker NR, McKiernan M. Modifications to the photosynthetic apparatus of higher plants in response to changes in the light environment. Biol J Linn Soc. 1988;34:193–203.
-
- Björkman O. Responses to different quantum flux densities. In: Lange OL, Nobel PS, Osmond CB, Ziegler H, editors. Encyclopedia of Plant Physiology: Physiological Plant Ecology I (New Series), Vol 12A. Berlin: Springer-Verlag; 1981. pp. 57–107.
-
- Coleman JS, McConnaughay KDM, Bazzaz FA. Elevated CO2 and plant nitrogen-use: is reduced tissue nitrogen concentration size-dependent? Oecologia. 1993;93:195–200. - PubMed
-
- Dalling MJ, Boland G, Wilson JH. Relationship between acid protease activity and redistribution of nitrogen during grain development in wheat. Aust J Plant Physiol. 1976;3:721–730.
LinkOut - more resources
Full Text Sources
