

Expression of tobacco carbonic anhydrase in the C4 dicot flaveria bidentis leads to increased leakiness of the bundle sheath and a defective CO2-concentrating mechanism
- PMID: 9662550
- PMCID: PMC34923
- DOI: 10.1104/pp.117.3.1071
Expression of tobacco carbonic anhydrase in the C4 dicot flaveria bidentis leads to increased leakiness of the bundle sheath and a defective CO2-concentrating mechanism
Abstract
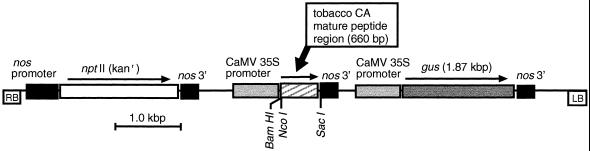
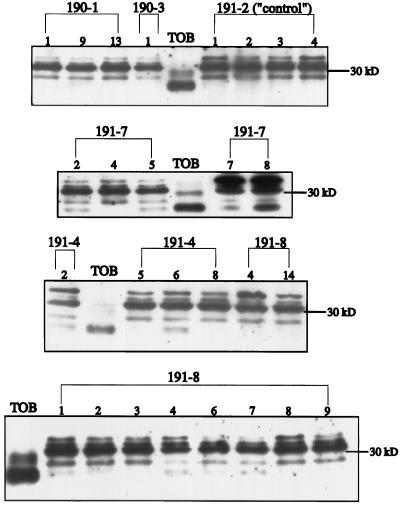
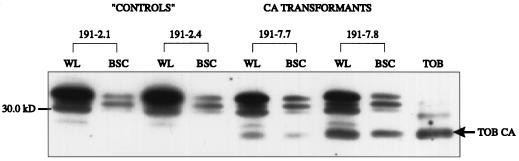
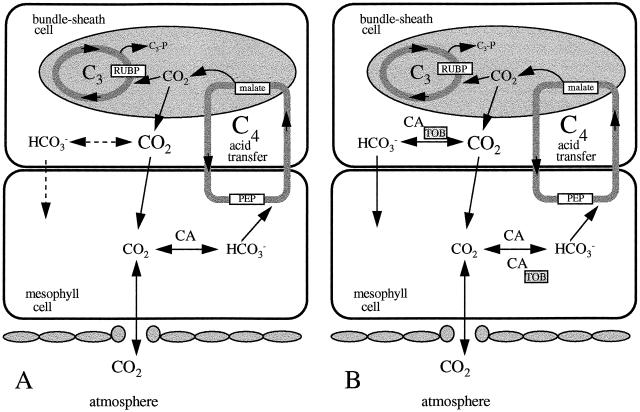
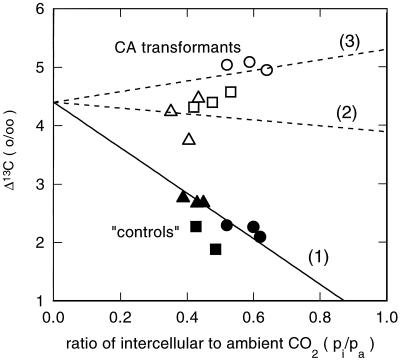
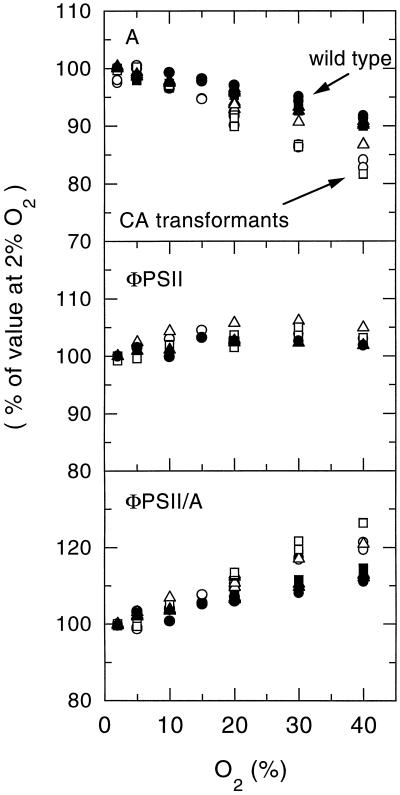
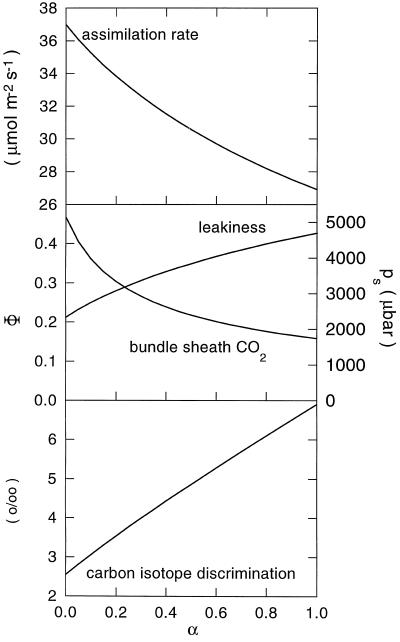
Flaveria bidentis (L.) Kuntze, a C4 dicot, was genetically transformed with a construct encoding the mature form of tobacco (Nicotiana tabacum L.) carbonic anhydrase (CA) under the control of a strong constitutive promoter. Expression of the tobacco CA was detected in transformant whole-leaf and bundle-sheath cell (bsc) extracts by immunoblot analysis. Whole-leaf extracts from two CA-transformed lines demonstrated 10% to 50% more CA activity on a ribulose-1,5-bisphosphate carboxylase/oxygenase-site basis than the extracts from transformed, nonexpressing control plants, whereas 3 to 5 times more activity was measured in CA transformant bsc extracts. This increased CA activity resulted in plants with moderately reduced rates of CO2 assimilation (A) and an appreciable increase in C isotope discrimination compared with the controls. With increasing O2 concentrations up to 40% (v/v), a greater inhibition of A was found for transformants than for wild-type plants; however, the quantum yield of photosystem II did not differ appreciably between these two groups over the O2 levels tested. The quantum yield of photosystem II-to-A ratio suggested that at higher O2 concentrations, the transformants had increased rates of photorespiration. Thus, the expression of active tobacco CA in the cytosol of F. bidentis bsc and mesophyll cells perturbed the C4 CO2-concentrating mechanism by increasing the permeability of the bsc to inorganic C and, thereby, decreasing the availability of CO2 for photosynthetic assimilation by ribulose-1,5-bisphosphate carboxylase/oxygenase.
Figures
Similar articles
-
Carbonic anhydrase and its influence on carbon isotope discrimination during C4 photosynthesis. Insights from antisense RNA in Flaveria bidentis.Plant Physiol. 2006 May;141(1):232-42. doi: 10.1104/pp.106.077776. Epub 2006 Mar 16. Plant Physiol. 2006. PMID: 16543411 Free PMC article.
-
Expressing an RbcS Antisense Gene in Transgenic Flaveria bidentis Leads to an Increased Quantum Requirement for CO2 Fixed in Photosystems I and II.Plant Physiol. 1997 Nov;115(3):1163-1174. doi: 10.1104/pp.115.3.1163. Plant Physiol. 1997. PMID: 12223865 Free PMC article.
-
Reduction of Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase by Antisense RNA in the C4 Plant Flaveria bidentis Leads to Reduced Assimilation Rates and Increased Carbon Isotope Discrimination.Plant Physiol. 1997 Feb;113(2):469-477. doi: 10.1104/pp.113.2.469. Plant Physiol. 1997. PMID: 12223620 Free PMC article.
-
Strategies for improving C4 photosynthesis.Curr Opin Plant Biol. 2016 Jun;31:125-34. doi: 10.1016/j.pbi.2016.04.003. Epub 2016 Apr 27. Curr Opin Plant Biol. 2016. PMID: 27127850 Review.
-
Bundle-sheath leakiness in C4 photosynthesis: a careful balancing act between CO2 concentration and assimilation.J Exp Bot. 2014 Jul;65(13):3443-57. doi: 10.1093/jxb/eru157. Epub 2014 Apr 22. J Exp Bot. 2014. PMID: 24755278 Review.
Cited by
-
Meeting the photosynthetic demand for inorganic carbon in an alga-invertebrate association: preferential use of CO2 by symbionts in the giant clam Tridacna gigas.Proc Biol Sci. 2000 Mar 7;267(1442):523-9. doi: 10.1098/rspb.2000.1031. Proc Biol Sci. 2000. PMID: 10737411 Free PMC article.
-
Expression of a CO2-permeable aquaporin enhances mesophyll conductance in the C4 species Setaria viridis.Elife. 2021 Nov 29;10:e70095. doi: 10.7554/eLife.70095. Elife. 2021. PMID: 34842138 Free PMC article.
-
Light and growth temperature alter carbon isotope discrimination and estimated bundle sheath leakiness in C4 grasses and dicots.Photosynth Res. 2007 Jan;91(1):47-58. doi: 10.1007/s11120-007-9136-6. Epub 2007 Feb 27. Photosynth Res. 2007. PMID: 17333508
-
The roles of carbonic anhydrases in photosynthetic CO(2) concentrating mechanisms.Photosynth Res. 2003;77(2-3):83-94. doi: 10.1023/A:1025821717773. Photosynth Res. 2003. PMID: 16228367
-
Internal conductance to CO(2) diffusion and C(18)OO discrimination in C(3) leaves.Plant Physiol. 2000 May;123(1):201-14. doi: 10.1104/pp.123.1.201. Plant Physiol. 2000. PMID: 10806237 Free PMC article.
References
-
- Berry J, Farquhar G (1978) The CO2 concentrating function of C4 photosynthesis: a biochemical model. In D Hall, J Coombs, TW Goodwin, eds, Proceedings of the Fourth International Congress on Photosynthesis. Biochemical Society, London, pp 119–131
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
