

doi: 10.1084/jem.188.2.341.
Immune pathogenesis of hepatocellular carcinoma
Affiliations
- PMID: 9670046
- PMCID: PMC2212453
- DOI: 10.1084/jem.188.2.341
Item in Clipboard
Immune pathogenesis of hepatocellular carcinoma
J Exp Med.
.
Abstract
Hepatocellular carcinoma (HCC) is a common complication of chronic hepatitis B virus (HBV) infection. The pathogenetic mechanisms potentially responsible for HCC during chronic HBV infection are not well defined. This study demonstrates that chronic immune-mediated liver cell injury triggers the development of HCC in the absence of viral transactivation, insertional mutagenesis, and genotoxic chemicals. These results strongly suggest that the immune response to HBV is both necessary and sufficient to cause liver cancer during chronic HBV infection, and that all other procarcinogenic events associated with HCC are probably dependent on this process.
Figures
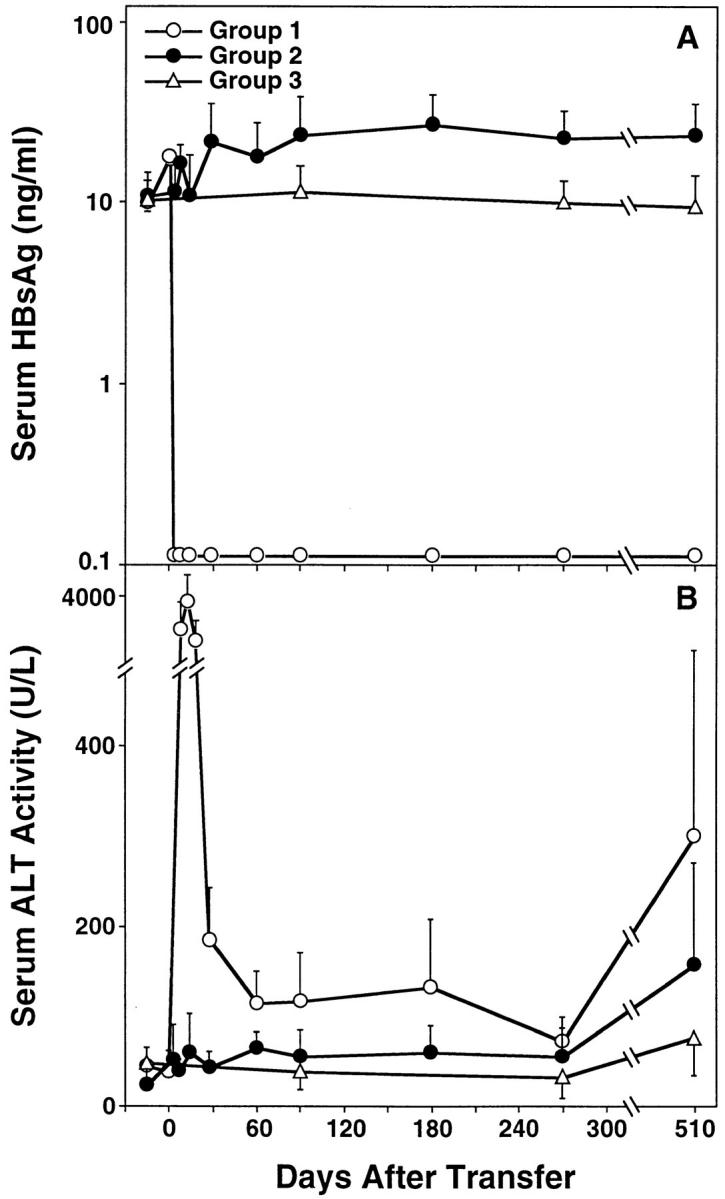
Kinetics of serum HBsAg (A) and ALT (B) in HBV transgenic mice. Group 1 mice (open circles; n = 9) were thymectomized, lethally irradiated, and reconstituted with bone marrow and spleen cells from syngeneic nontransgenic donors that had been previously immunized with recombinant vaccinia virus HBs-vac. Similarly treated group 2 transgenic control animals (filled circles; n = 9) were reconstituted with bone marrow and spleen cells from immunologically tolerant transgenic donors that had been injected with saline. All results were compared with group 3 unmanipulated age- and sex-matched transgenic mice (open triangles; n = 10). Adoptive transfer of splenocytes was performed on day 0. Results were expressed as mean units per liter ± SEM of sALT activity, and differences between experimental and control groups were assessed for statistical significance by Student's t test.
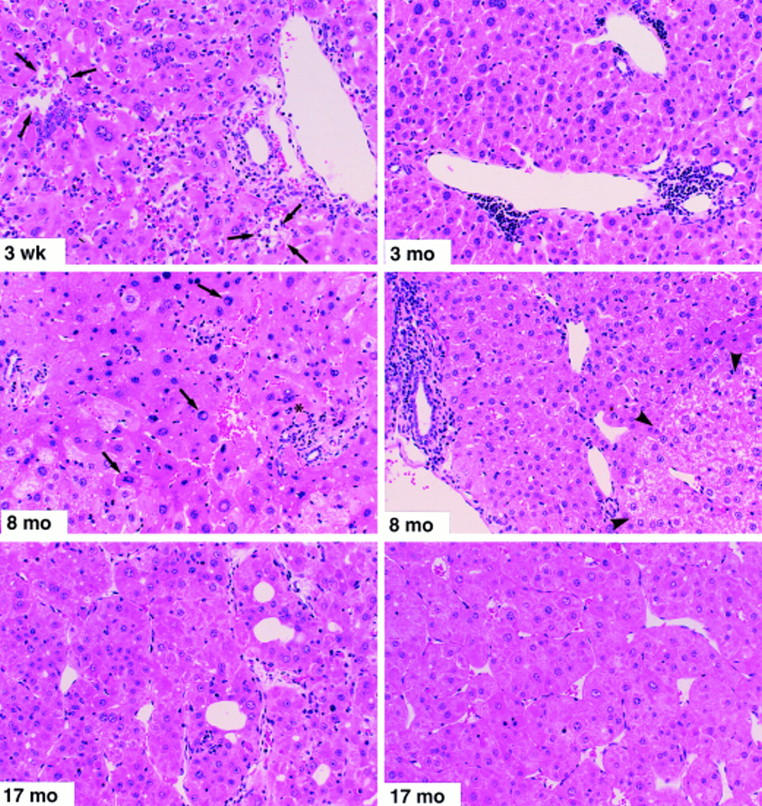
Histopathological features of prolonged chronic immune-mediated hepatitis in HBV transgenic mice (group 1). Mice were killed by cervical dislocation at the indicated time points. The 3-wk specimen illustrates portal and parenchymal inflammatory infiltrates with focal hepatocellular dropout (arrows) and hepatocellular apoptosis (asterisk). The 3-mo specimen displays lymphocytic portal infiltrates and moderate disarray of the lobular architecture. In addition, several lobular inflammatory foci were observed at this time point (not shown). The 8-mo specimen (left) illustrates marked lobular disarray, small focal lobular inflammatory infiltrates associated with degenerating hepatocytes, acidophil bodies (asterisk), and marked variation in the size and shape of many hepatocytes, some of which display nuclear pseudoinclusions (arrows). The other 8-mo specimen (right) illustrates a portal inflammatory infiltrate (at the left of the photo) with piecemeal necrosis of the limiting plate, and a glycogen-rich altered hepatic focus (arrowheads) containing numerous hepatocytes with prominent nucleoli (features traditionally associated with preneoplastic foci). Both of the 17-mo specimens, taken from large (>15 mm diameter) tumors from different animals, illustrate the classical histological features of trabecular HCC consisting of relatively well-differentiated hepatocytes with granular eosinophilic cytoplasm, arranged in nests and cords several cells thick that display increased mitotic activity, compress the adjacent hepatic parenchyma, and display areas of hemorrhage and necrosis (not shown). Hematoxylin and eosin; original magnification: ×200.
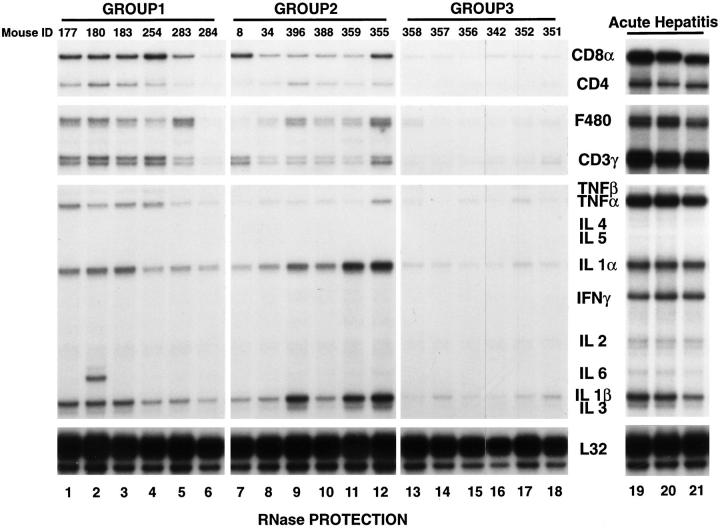
Intrahepatic T cell and cytokine profiles in HBV transgenic mice. Total RNA was extracted from nontumorous liver tissue of representative mice from each group 17 mo after adoptive transfer (groups 1 and 2) and from unmanipulated age-matched controls (group 3), and subjected to RNase protection analysis to monitor the expression of CD3γ, CD4, CD8α, F480, and a panel of inflammatory cytokines. Lanes 19–21 contain total liver RNA from three HBV transgenic mice that were injected with an HBsAg-specific CTL clone which induced a severe transient acute hepatitis in transgenic recipients (references and 28). The ribosomal protein light 32 (L32) RNA was used to normalize the amount of RNA loaded in each lane.
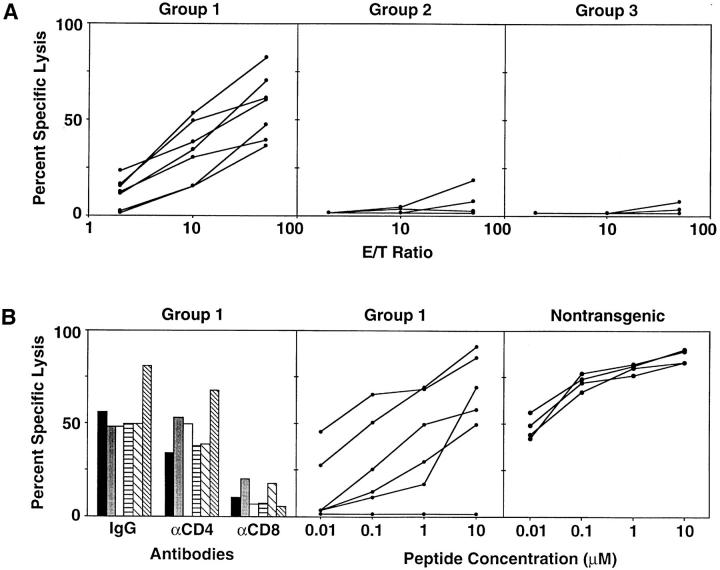
(A) In vitro cytotoxic activity of splenocytes from group 1, 2, and 3 mice at the time of autopsy. Splenocytes from individual mice were stimulated in vitro every 7 d with irradiated P815 cells that stably express the HBV envelope proteins (P815preS1 cells). 2 wk later, the cytolytic activity of the resultant T cell lines was assessed in a standard 4-h cytotoxicity assay with 51Cr-labeled P815 cells and P815 transfectants that express the HBV small envelope polypeptide (P815-S) at the E/T ratios shown. Each line represents the cytolytic activity detectable in an individual mouse. (B) Characteristics of in vitro cytotoxic activity of splenocytes from group 1 mice and HBs-vac–primed nontransgenic mice. Left, The influence of anti-CD4 and anti-CD8 Abs on the HBsAg-specific cytolytic activity of group 1 spleen cells that had been stimulated for an additional 1 wk and tested for cytolytic activity against P815 and P815-S target cells at an E/T ratio of 50:1 in the presence of a rat mAb specific for mouse CD4 or CD8 or a control Ab. Columns with the same pattern indicate cytotoxic activity of splenocytes from the same mouse. Middle, Cytolytic activity of group 1 spleen cells against P815 cells that had been pulsed either with the indicated concentrations of a synthetic peptide representing residues 28–39 of HBsAg or with media at an E/T ratio of 50:1. Right, Cytolytic activity of similarly stimulated splenocytes from HBs-vac–primed nontransgenic B10D2 (H-2d) mice identical to those used as group 1 donors. Middle and right, Each line represents the cytolytic activity detectable in an individual mouse.
References
-
- Chisari FV, Ferrari C. Hepatitis B virus immunopathogenesis. Annu Rev Immunol. 1995;13:29–60. - PubMed
-
- Tsai SL, Chen PJ, Lai MY, Yang PM, Sung JL, Huang JH, Hwang LH, Chang TH, Chen DS. Acute exacerbations of chronic type B hepatitis are accompanied by increased T cell responses to hepatitis B core and e antigens. Implications for hepatitis B e antigen seroconversion. J Clin Invest. 1992;89:87–96. - PMC - PubMed
-
- Colombo M, de Franchis R, Del Ninno E, Sangiovanni A, De Fazio C, Tommasini M, Donato MF, Piva A, Di Carlo V, Dioguardi N. Hepatocellular carcinoma in Italian patients with cirrhosis. N Engl J Med. 1991;325:675–680. - PubMed
-
- Okuda K. Hepatocellular carcinoma: recent progress. Hepatology. 1992;15:948–963. - PubMed
-
- Ikeda K, Saitoh S, Koida I, Arase Y, Tsubota A, Chayama K, Kumada H, Kawanishi M. A multivariate analysis of risk factors for hepatocellular carcinogenesis: a prospective observation of 795 patients with viral and alcoholic cirrhosis. Hepatology. 1993;18:47–53. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
