

Host resistance to intracellular infection: mutation of natural resistance-associated macrophage protein 1 (Nramp1) impairs phagosomal acidification
- PMID: 9670047
- PMCID: PMC2212455
- DOI: 10.1084/jem.188.2.351
Host resistance to intracellular infection: mutation of natural resistance-associated macrophage protein 1 (Nramp1) impairs phagosomal acidification
Abstract
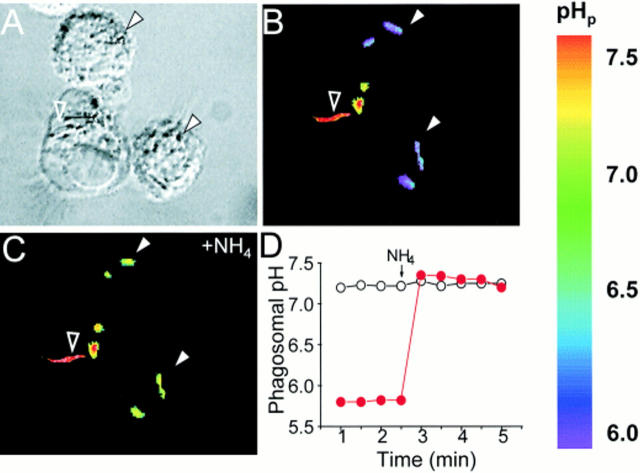
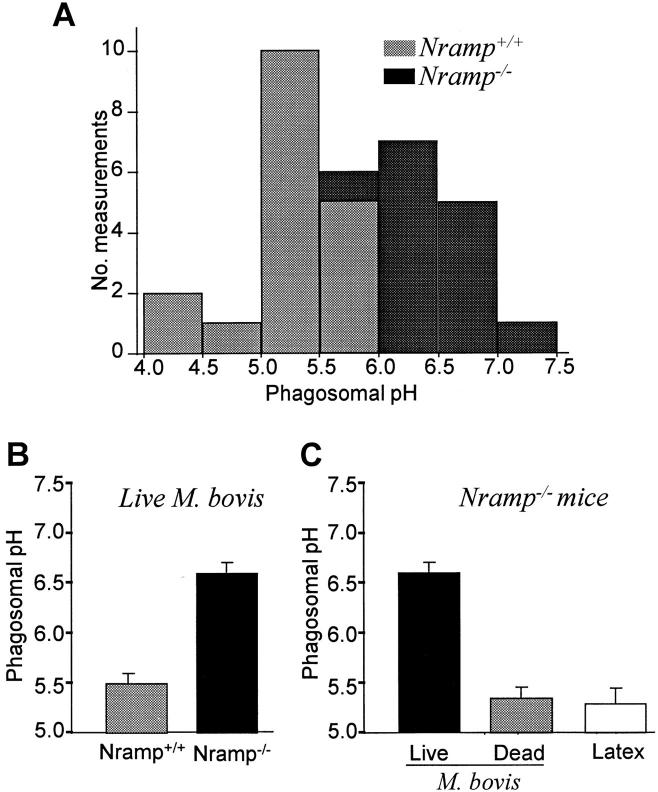
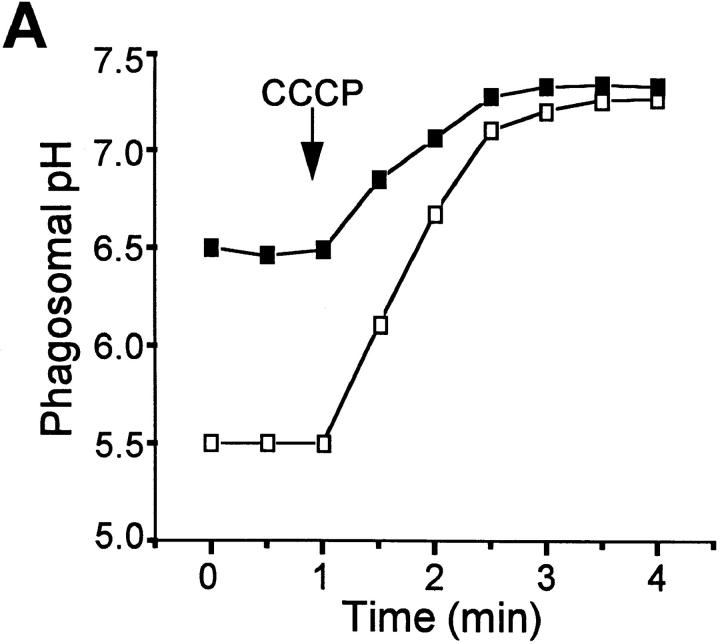
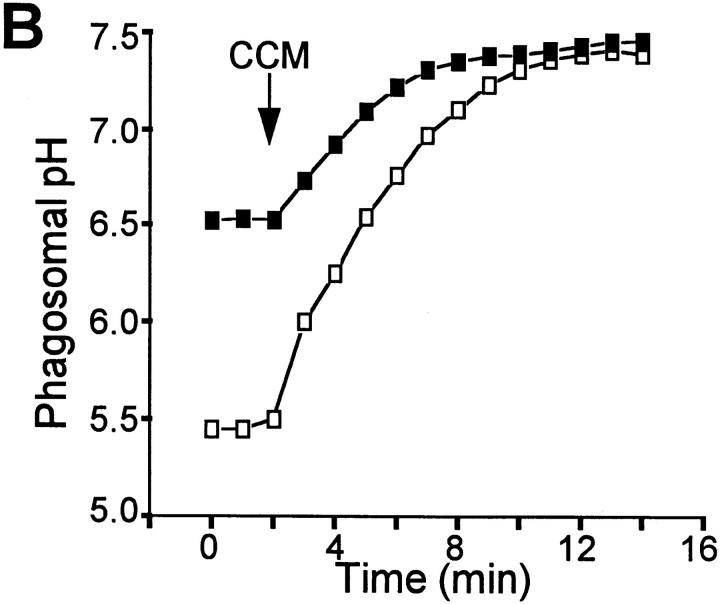
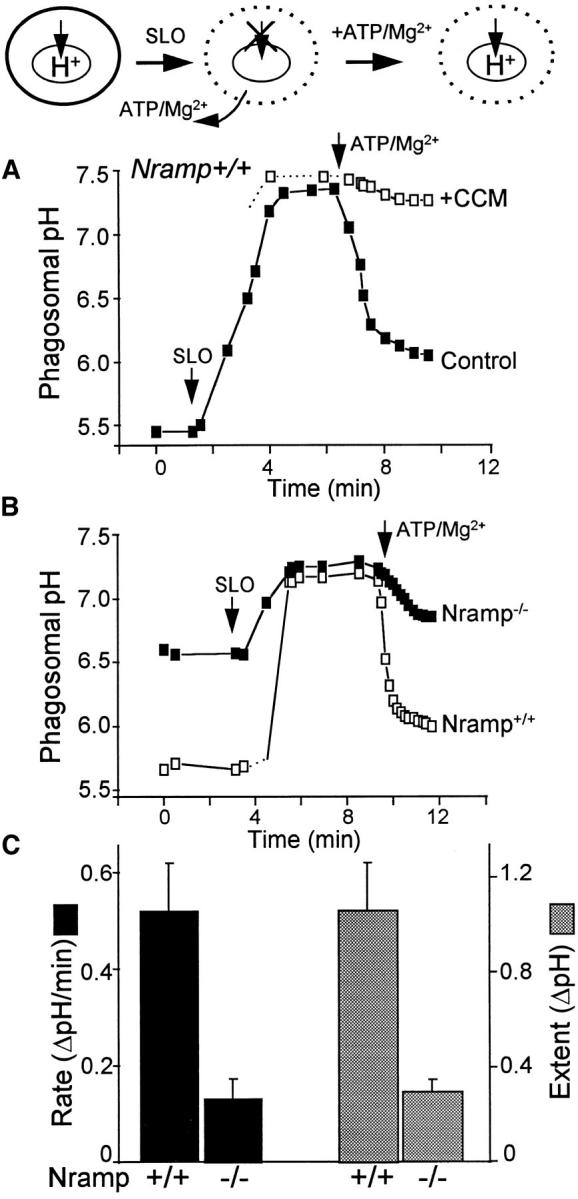
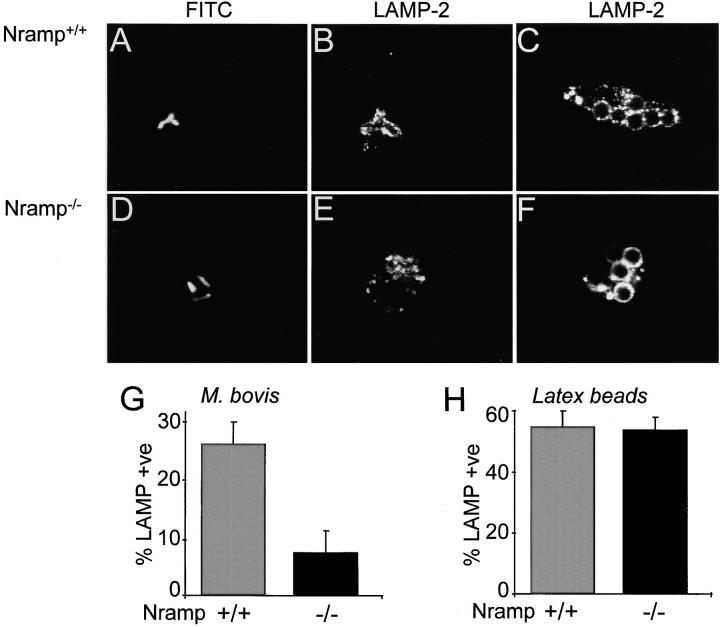
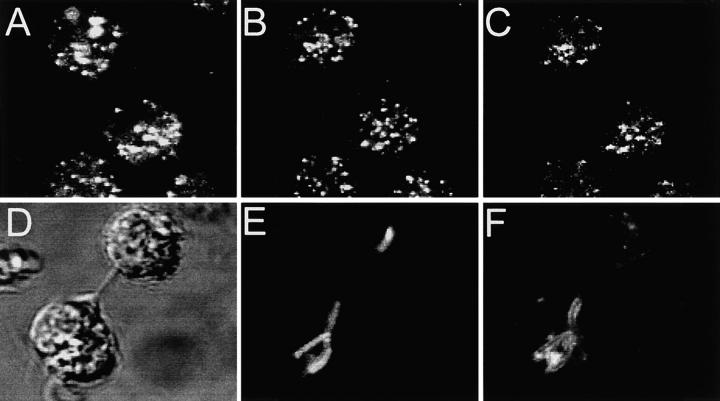
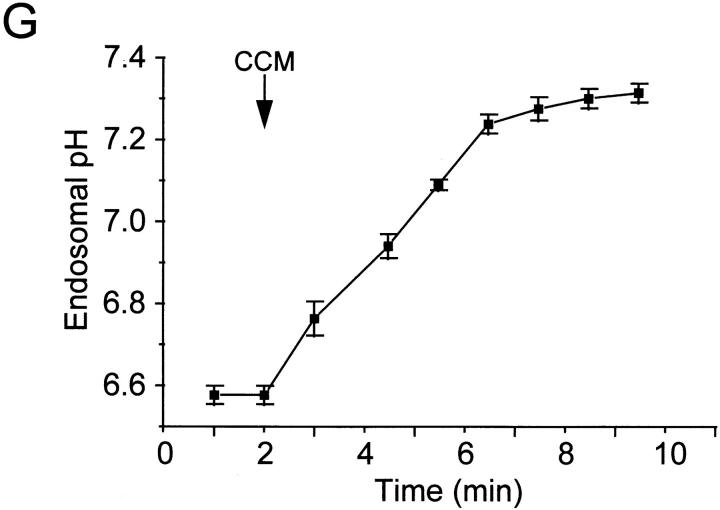
The mechanisms underlying the survival of intracellular parasites such as mycobacteria in host macrophages remain poorly understood. In mice, mutations at the Nramp1 gene (for natural resistance-associated macrophage protein), cause susceptibility to mycobacterial infections. Nramp1 encodes an integral membrane protein that is recruited to the phagosome membrane in infected macrophages. In this study, we used microfluorescence ratio imaging of macrophages from wild-type and Nramp1 mutant mice to analyze the effect of loss of Nramp1 function on the properties of phagosomes containing inert particles or live mycobacteria. The pH of phagosomes containing live Mycobacterium bovis was significantly more acidic in Nramp1- expressing macrophages than in mutant cells (pH 5.5 +/- 0.06 versus pH 6.6 +/- 0.05, respectively; P <0.005). The enhanced acidification could not be accounted for by differences in proton consumption during dismutation of superoxide, phagosomal buffering power, counterion conductance, or in the rate of proton "leak", as these were found to be comparable in wild-type and Nramp1-deficient macrophages. Rather, after ingestion of live mycobacteria, Nramp1-expressing cells exhibited increased concanamycin-sensitive H+ pumping across the phagosomal membrane. This was associated with an enhanced ability of phagosomes to fuse with vacuolar-type ATPase-containing late endosomes and/or lysosomes. This effect was restricted to live M. bovis and was not seen in phagosomes containing dead M. bovis or latex beads. These data support the notion that Nramp1 affects intracellular mycobacterial replication by modulating phagosomal pH, suggesting that Nramp1 plays a central role in this process.
Figures
References
-
- Young P. White House to expand response to infectious diseases. Am Soc Microbiol News. 1996;62:450–451.
-
- Berkelman RL, Bryan RT, Osterholm MT, Leduc JW, Hughes JM. Infectious disease surveillance: a crumbling foundation. Science. 1996;264:368–370. - PubMed
-
- Bloom BR. Tuberculosis: back to a frightening future. Nature. 1992;358:538–540. - PubMed
-
- Kaye K, Frieden TR. Tuberculosis control: the relevance of classic principles in an era of acquired immunodeficiency syndrome and multidrug resistance. Epidemiologic Rev. 1996;18:52–63. - PubMed
-
- Davies J. Inactivation of antibiotics and the dissemination of resistance genes. Science. 1994;264:375–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
