

A potential role for U2AF-SAP 155 interactions in recruiting U2 snRNP to the branch site
- PMID: 9671485
- PMCID: PMC109061
- DOI: 10.1128/MCB.18.8.4752
A potential role for U2AF-SAP 155 interactions in recruiting U2 snRNP to the branch site
Abstract
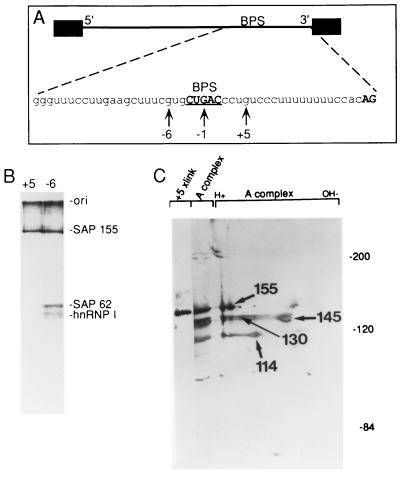
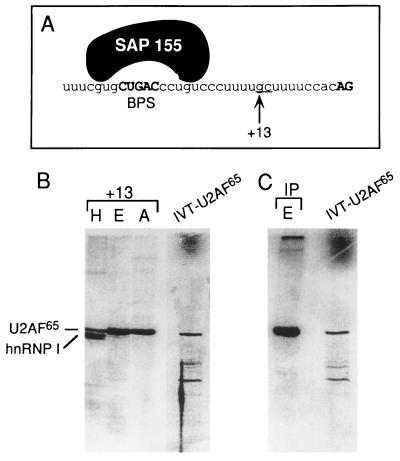
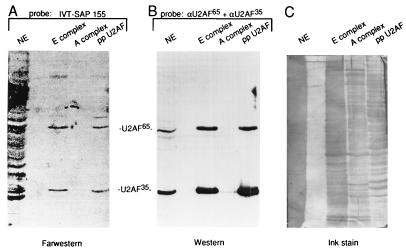
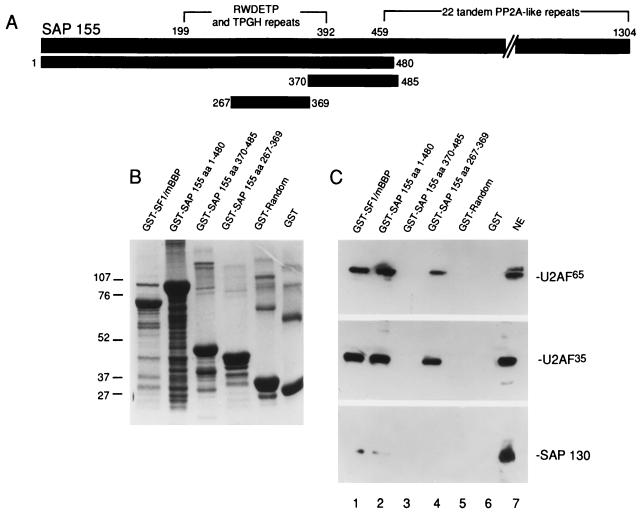
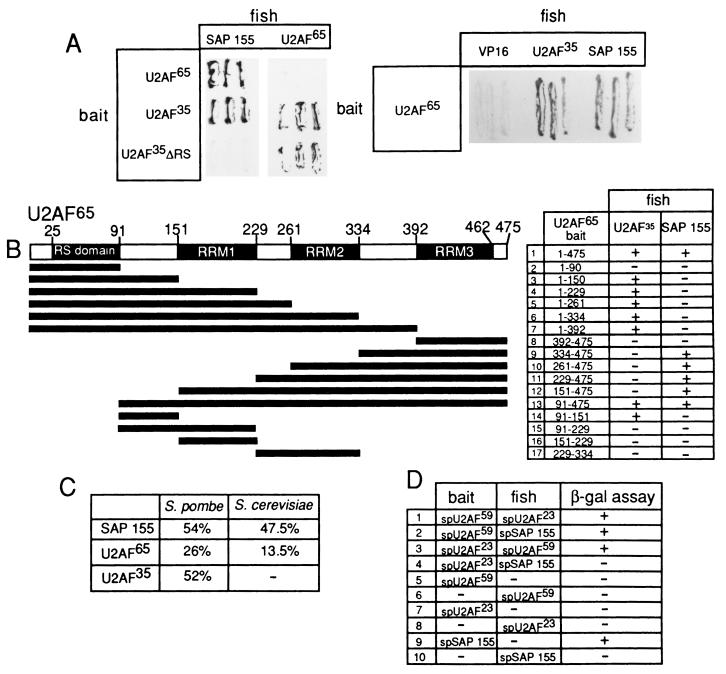
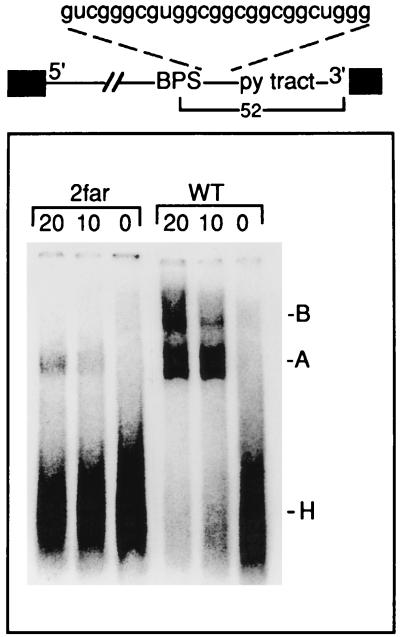
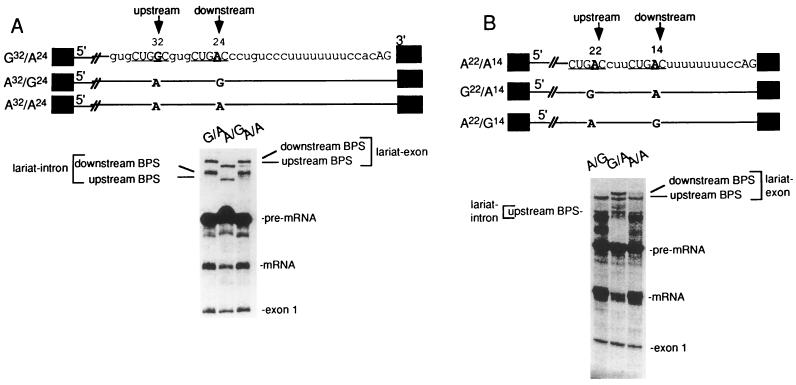
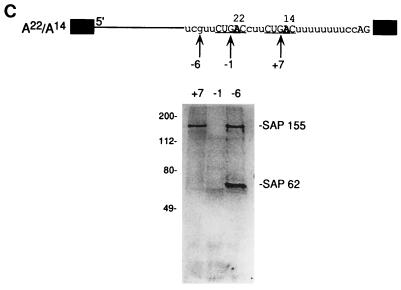
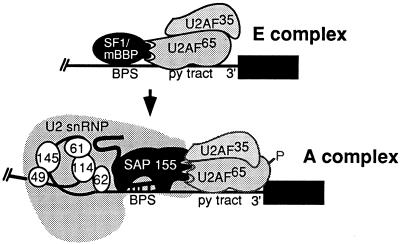
Base pairing between U2 snRNA and the branchpoint sequence (BPS) is essential for pre-mRNA splicing. Because the metazoan BPS is short and highly degenerate, this interaction alone is insufficient for specific binding of U2 snRNP. The splicing factor U2AF binds to the pyrimidine tract at the 3' splice site in the earliest spliceosomal complex, E, and is essential for U2 snRNP binding in the spliceosomal complex A. We show that the U2 snRNP protein SAP 155 UV cross-links to pre-mRNA on both sides of the BPS in the A complex. SAP 155's downstream cross-linking site is immediately adjacent to the U2AF binding site, and the two proteins interact directly in protein-protein interaction assays. Using UV cross-linking, together with functional analyses of pre-mRNAs containing duplicated BPSs, we show a direct correlation between BPS selection and UV cross-linking of SAP 155 on both sides of the BPS. Together, our data are consistent with a model in which U2AF binds to the pyrimidine tract in the E complex and then interacts with SAP 155 to recruit U2 snRNP to the BPS.
Figures
References
-
- Abovich N, Liao X C, Rosbash M. The yeast MUD2 protein: an interaction with PRP11 defines a bridge between commitment complexes and U2 snRNP addition. Genes Dev. 1994;8:843–854. - PubMed
-
- Abovich N, Rosbash M. Cross-intron bridging interactions in the yeast commitment complex are conserved in mammals. Cell. 1997;89:403–412. - PubMed
-
- Bennett M, Reed R. Correspondence between a mammalian spliceosome component and an essential yeast splicing factor. Science. 1993;262:105–108. - PubMed
-
- Bennett M, Michaud S, Kingston J, Reed R. Protein components specifically associated with prespliceosome and spliceosome complexes. Genes Dev. 1992;6:1986–2000. - PubMed
-
- Berglund J A, Chua K, Abovich N, Reed R, Rosbash M. The splicing factor BBP interacts specifically with the pre-mRNA branchpoint sequence UACUAAC. Cell. 1997;89:781–787. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous