

doi: 10.1073/pnas.95.15.8538.
Mammalian mediator of transcriptional regulation and its possible role as an end-point of signal transduction pathways
Affiliations
- PMID: 9671713
- PMCID: PMC21111
- DOI: 10.1073/pnas.95.15.8538
Item in Clipboard
Mammalian mediator of transcriptional regulation and its possible role as an end-point of signal transduction pathways
Proc Natl Acad Sci U S A.
.
Abstract
A multiprotein complex isolated from murine cells is identified as a counterpart of the yeast Mediator of transcriptional regulation on the basis of the following: homologs of two subunits of yeast Mediator, Srb7 and Med7, copurify with the complex; peptide sequencing reveals, in addition, homologs of the yeast Mediator subunits Rgr1 and Med6; as with yeast Mediator, the mouse complex binds to the RNA polymerase II C-terminal domain (CTD) and stimulates phosphorylation of the CTD by TFIIH. Peptide sequencing also identifies a component of mouse Mediator as a relative of Ring-3 protein, a mitogen-activated nuclear protein kinase, raising the possibility of Mediator as an end point of signal transduction pathways.
Figures
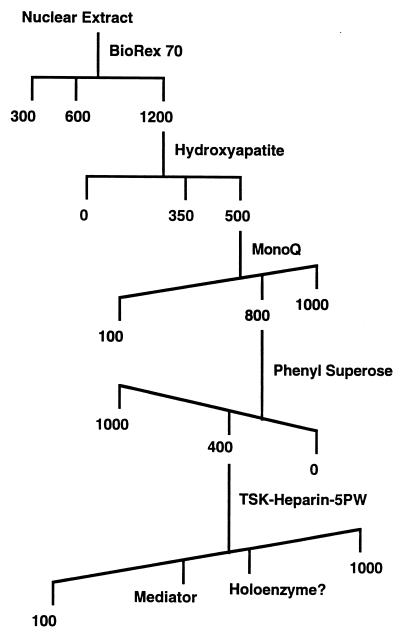
Mouse Mediator fractionation scheme. Numbers indicate concentrations (mM) of salts used for elution.
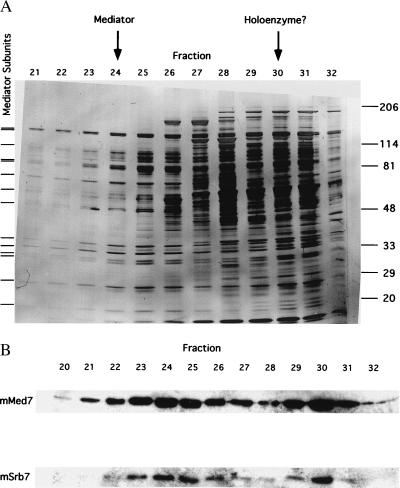
Elution profiles from last step of mouse Mediator purification. (A) SDS/PAGE of TSK-Heparin-5PW fractions. Proteins were revealed by silver staining. Tick marks at left indicate positions of bands that coelute and coimmunoprecipitate (Fig. 3) and that are therefore attributed to Mediator subunits. Positions of bands due to size markers (kDa) are indicated at the right. Peak fractions of mSrb7 and mMed7 in the immunoblot (B Lower), attributed to Mediator and to possible RNA polymerase II–Mediator complex (“holoenzyme”), are indicated by arrows at the top. (B) Immunoblot of TSK-Heparin-5PW fractions with antibodies against hSrb7 and hMed7. Only the regions of the blots containing mSrb7 and mMed7 are shown.
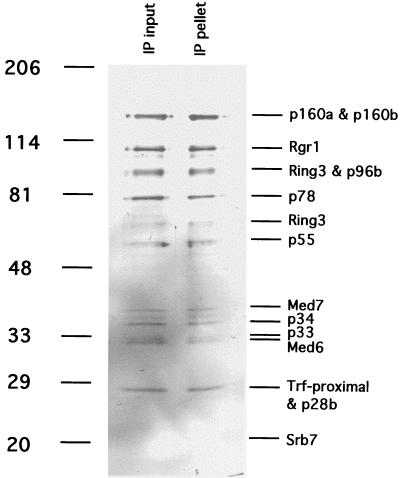
Immunoprecipitation of mouse Mediator with antibodies against hSrb7. TSK-Heparin-5PW fraction 24 (50 μl) was immunoprecipitated with 100 μl of protein A-Sepharose-purified anti-hSrb7 antibodies crosslinked to 50 μl of Sepharose beads as described (5). After incubation for 4 h at 4°C in 25 mM Tris acetate, pH 7.8/200 mM potassium acetate/0.1 mM DTT/1 mM EDTA/10% glycerol/protease inhibitors, the beads were washed twice with the same buffer containing 500 mM potassium acetate/0.2% Nonidet P-40. Immunoprecipitated proteins (“IP pellet”) were eluted with 5 M urea twice for 10 min at room temperature, precipitated with trichloroacetic acid, analyzed alongside the starting fraction (“IP input,” 20 μl, precipitated with trichloroacetic acid) by electrophoresis in an 8–12% gradient SDS-polyacrylamide gel, and revealed by silver staining. Positions of bands due to size markers (kDa) are indicated at the left. Mediator subunits are identified at the right.

Sequence alignments. (A) Mouse and C. elegans Rgr1 homologs. Sequence analysis of the p110 component of mouse Mediator yielded two peptides (underlined) matching the sequence expected from mouse EST AA562494 translated in the +2 frame. The DNA sequence of EST AA562494 partially overlapped that of EST AA204093, and fusion of the two DNA sequences and translation generated the 233-residue mouse sequence shown. A blast search with this sequence revealed the homology with C. elegans Rgr1 shown. A third peptide from p110 was mapped to a region of the 233-residue mouse sequence (indicated by asterisks) on the basis of its experimental mass (m/z = 2122.097), which was in excellent agreement with the calculated monoisotopic mass of the expected peptide [(MH+) = 2122.123, Δ = 0.026 Da (12 ppm)]. (B) Yeast Rgr1 and C. elegans Rgr1 homolog. (C) Human homolog of the p28a component of mouse Mediator and Drosophila Trf-proximal. Peptide sequences obtained from p28a are underlined. A difference between human and mouse sequences is indicated by an asterisk.
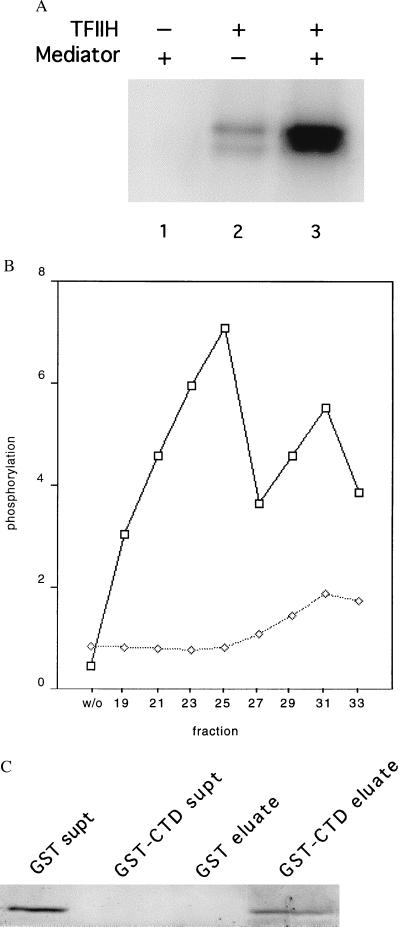
Physical and functional interaction of mouse Mediator with the RNA polymerase II CTD. (A) Stimulation by mouse Mediator of CTD phosphorylation by TFIIH. Recombinant His-tagged yeast CTD (50 ng) was incubated with rat TFIIH (15 ng) and mouse Mediator (TSK-Heparin-5PW fraction 25, 100 ng) in 15 μl of 20 mM Hepes, pH 7.6/110 mM potassium acetate/5 μM (0.3 μCi) [γ-32P]ATP/7.5 mM magnesium acetate/2 mM DTT for 30 min at 23°C, followed by electrophoresis in an SDS-polyacrylamide gel and autoradiography. (B) Profile of stimulatory activity shown in A across the TSK-Heparin-5PW column (1 μl of each fraction, corresponding to 100 ng of fraction 25, or control without fraction, designated “w/o”), measured in the presence (upper curve) or absence (lower curve) of added TFIIH. Intensities of bands determined with a PhosphorImager are plotted in arbitrary units on the abscissa. (C) Binding of mouse Mediator to GST-CTD. Glutathione-agarose (30 μl of a 50% slurry, Sigma) was incubated with GST (50 μg) or GST-CTD (50 μg) for 1 h at 4°C with gentle agitation. The beads were washed three times with buffer E(600) (20 mM Tris⋅Cl, pH 7.9/1 mM MgCl2/10 μM ZnCl2/0.2 mM EDTA/10% glycerol/protease inhibitors; mM potassium acetate in parenthesis) containing 0.02% Nonidet P-40, and twice in buffer E(200) containing 0.01% Nonidet P-40. The beads then were incubated with Mediator (TSK-Heparin-5PW fraction 25, 1.5 μg) in 100 μl of buffer E(200) containing 0.01% Nonidet P-40 for 2 h at 4°C with gentle agitation, followed by three washes with buffer E(200) containing 0.1% Nonidet P-40 and elution with 35 μl of 10 mM reduced glutathione in 50 mM Tris acetate, pH 7.8, for 10 min at 4°C with gentle agitation. The supernatants from the initial binding of Mediator, concentrated by trichloroacetic acid precipitation (“GST supt,” “GST-CTD supt”), and the entire eluates were analyzed by SDS/PAGE and immunoblotting with anti-mMed7 antibodies.
References
-
- Stringer K F, Ingles C J, Greenblatt J. Nature (London) 1990;345:783–786. - PubMed
-
- Flanagan P M, Kelleher R J, III, Sayre M H, Tschochner H, Kornberg R D. Nature (London) 1991;350:436–438. - PubMed
-
- Pugh B F, Tjian R. Cell. 1990;61:1187–1197. - PubMed
-
- Zhou Q, Liberman P M, Boyer T G, Berk A J. Genes Dev. 1992;6:1964–1974. - PubMed
-
- Kim Y J, Bjorklund S, Li Y, Sayre M H, Kornberg R D. Cell. 1994;77:599–608. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
