

The exocyclic groups of DNA modulate the affinity and positioning of the histone octamer
- PMID: 9671714
- PMCID: PMC21112
- DOI: 10.1073/pnas.95.15.8544
The exocyclic groups of DNA modulate the affinity and positioning of the histone octamer
Abstract
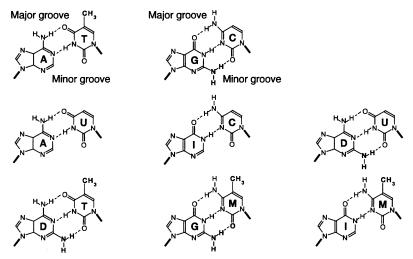
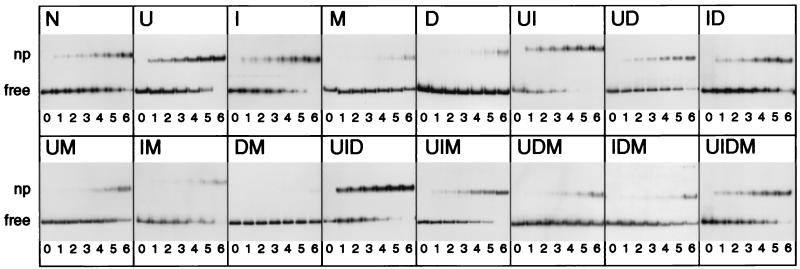
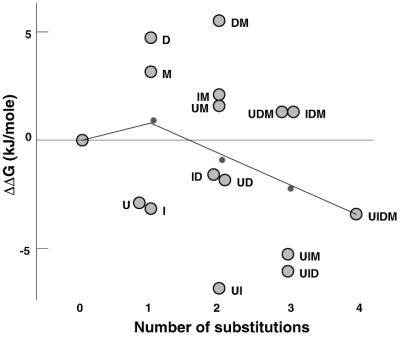
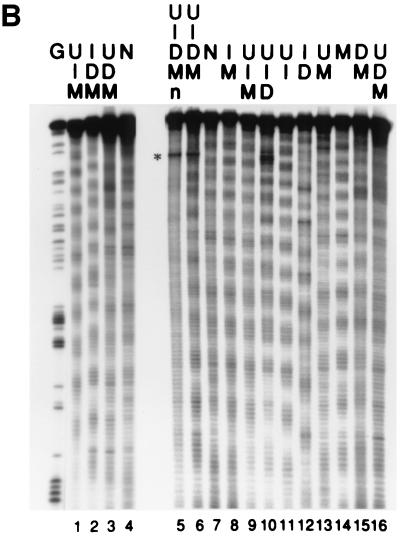
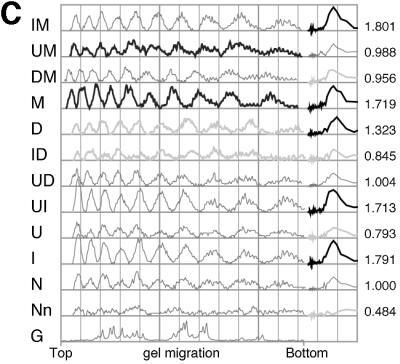
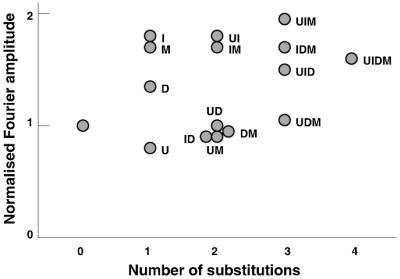
To investigate the nature of the chemical determinants in DNA required for nonspecific binding and bending by proteins we have created a novel DNA in which inosine-5-methylcytosine and 2, 6-diaminopurine-uracil base pairs are substituted for normal base pairs in a defined DNA sequence. This procedure completely switches the patterns of the base pair H bonding and attachment of exocyclic groups. We show that this DNA binds a histone octamer more tightly than normal DNA but, surprisingly, does not alter the orientation of the sequence on the surface of the protein. However, in general, the addition or removal of DNA exocyclic groups reduces or increases, respectively, the affinity for the histone octamer. The average incremental change in binding energy for a single exocyclic group is approximately 40 J/mol. The orientation of the DNA in core nucleosomes also is sensitive to the number and nature of the exocyclic groups present. Notably, substitution with the naturally occurring cytosine analogue, 5-methylcytosine, shifts the preferred rotational position by 3 bp, whereas incorporating 2,6-diaminopurine shifts it 2 bp in the opposite direction. These manipulations potentially would alter the accessibility of a protein recognition sequence on the surface of the histone octamer. We propose that exocyclic groups impose steric constraints on protein-induced DNA wrapping and are also important in determining the orientation of DNA on a protein surface. In addition, we consider the implications of the selection of A-T and G-C base pairs in natural DNA.
Figures

Similar articles
-
DNA recognition and nucleosome organization.Biopolymers. 1997;44(4):423-33. doi: 10.1002/(SICI)1097-0282(1997)44:4<423::AID-BIP6>3.0.CO;2-M. Biopolymers. 1997. PMID: 9782778 Review.
-
Relative affinities of DNA sequences for the histone octamer depend strongly upon both the temperature and octamer concentration.Biochemistry. 2005 Nov 1;44(43):14329-34. doi: 10.1021/bi050915w. Biochemistry. 2005. PMID: 16245949
-
Formation, stability and core histone positioning of nucleosomes reassembled on bent and other nucleosome-derived DNA.J Mol Biol. 1989 May 5;207(1):183-92. doi: 10.1016/0022-2836(89)90449-x. J Mol Biol. 1989. PMID: 2738923
-
Sequence-specific positioning of core histones on an 860 base-pair DNA. Experiment and theory.J Mol Biol. 1987 May 5;195(1):143-73. doi: 10.1016/0022-2836(87)90333-0. J Mol Biol. 1987. PMID: 3656408
-
The bending of DNA in nucleosomes and its wider implications.Philos Trans R Soc Lond B Biol Sci. 1987 Dec 15;317(1187):537-61. doi: 10.1098/rstb.1987.0080. Philos Trans R Soc Lond B Biol Sci. 1987. PMID: 2894688 Review.
Cited by
-
Exocyclic groups in the minor groove influence the backbone conformation of DNA.Nucleic Acids Res. 2001 Dec 15;29(24):5036-43. doi: 10.1093/nar/29.24.5036. Nucleic Acids Res. 2001. PMID: 11812834 Free PMC article.
-
Protein Engineering of Multi-Modular Transcription Factor Alcohol Dehydrogenase Repressor 1 (Adr1p), a Tool for Dissecting In Vitro Transcription Activation.Biomolecules. 2019 Sep 17;9(9):497. doi: 10.3390/biom9090497. Biomolecules. 2019. PMID: 31533362 Free PMC article.
-
Dissecting direct and indirect readout of cAMP receptor protein DNA binding using an inosine and 2,6-diaminopurine in vitro selection system.Nucleic Acids Res. 2008 Aug;36(14):4797-807. doi: 10.1093/nar/gkn452. Epub 2008 Jul 24. Nucleic Acids Res. 2008. PMID: 18653536 Free PMC article.
-
High-resolution biophysical analysis of the dynamics of nucleosome formation.Sci Rep. 2016 Jun 6;6:27337. doi: 10.1038/srep27337. Sci Rep. 2016. PMID: 27263658 Free PMC article.
-
Hepatitis delta antigen requires a flexible quasi-double-stranded RNA structure to bind and condense hepatitis delta virus RNA in a ribonucleoprotein complex.J Virol. 2014 Jul;88(13):7402-11. doi: 10.1128/JVI.00443-14. Epub 2014 Apr 16. J Virol. 2014. PMID: 24741096 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
