

doi: 10.1073/pnas.95.15.9031.
Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America
Affiliations
- PMID: 9671799
- PMCID: PMC21197
- DOI: 10.1073/pnas.95.15.9031
Item in Clipboard
Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America
Proc Natl Acad Sci U S A.
.
Abstract
Epidermal changes caused by a chytridiomycete fungus (Chytridiomycota; Chytridiales) were found in sick and dead adult anurans collected from montane rain forests in Queensland (Australia) and Panama during mass mortality events associated with significant population declines. We also have found this new disease associated with morbidity and mortality in wild and captive anurans from additional locations in Australia and Central America. This is the first report of parasitism of a vertebrate by a member of the phylum Chytridiomycota. Experimental data support the conclusion that cutaneous chytridiomycosis is a fatal disease of anurans, and we hypothesize that it is the proximate cause of these recent amphibian declines.
Figures
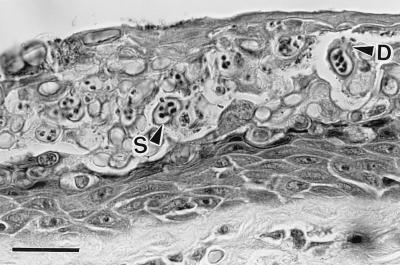
Histological section of severely infected digital skin of a wild frog, Litoria caerulea, from Queensland, Australia. The stratum corneum is markedly thickened because of a massive infection by a chytrid parasite. Thickness of normal stratum corneum is 2–5 μm, but here it is about 60 μm. This section contains a mass of intracellular sporangia (S) and developing sporangia. The mature sporangia are 12–20 μm (n = 25) in diameter, have refractile walls (0.5- to 2.0-μm thick), and contain zoospores. Many sporangia have discharged all zoospores. Zoospores are released through discharge tubes (D). Note the absence of an inflammatory cell reaction in the dermis and epidermis. (Bar = 30 μm.)
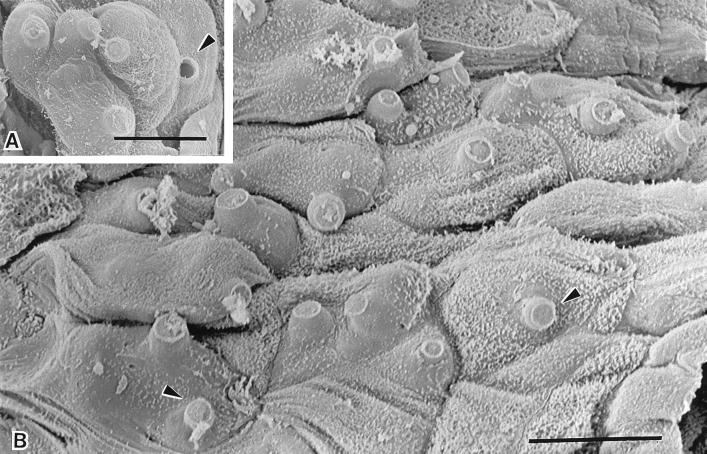
Scanning electron micrographs of infected digital skin of a wild frog, Litoria lesueuri, from Queensland, Australia, which died with cutaneous chytridiomycosis. (A) A cluster of mature sporangia are evident within the cells of the epidermis. The discharge tubes of the sporangia are visible projecting outwards from the cell surface. The plug of material within the discharge tube has been released in one of the sporangia (arrowhead). (Bar = 10 μm.) (B) Most of the cells in this field of the superficial layer of the epidermis contain sporangia, as evident by the bulging surfaces and the protrusion of unopened discharge tubes through infected cells (arrowheads). (Bar = 10 μm.)

Transmission electron micrographs of the zoospores found within fungal sporangia in the epidermis of naturally infected amphibians from Panama (Eleutherodactylus emcelae, A and C) and from Australia (Litoria caerulea, B and D). (A) Longitudinal section through a zoospore. The ribosomal area (Rb) is surrounded by a single cisterna of endoplasmic reticulum and bounded by a nucleus (N), mitochondria (Mi), and a microbody–lipid globule complex (asterisk). A single flagellum (Fl) connects to the posterior of the zoospore. (Bar = 1 μm.) (B) A transverse–oblique section through the anterior aspect of a zoospore demonstrates the unusual microbody–lipid globule complex. The microbody (asterisk) lies adjacent to four lipid globules (L) in this section. There is no evidence of a cisterna bounding these lipid globules. Rb, ribosomal area. (Bar = 1 μm.) (C) Transverse section through the anterior aspect of a zoospore. Note the discoidal cristae within mitochondria (Mi). The ribosomal area is bounded by the microbody–lipid globule complex (asterisk) and the mitochondria. No cisterna bound the lipid globules in this section. (Bar = 0.5 μm.) (D) A longitudinal–oblique section through a zoospore in the epidermis of an Australian amphibian for comparison with A. (Bar = 0.75 μm.)
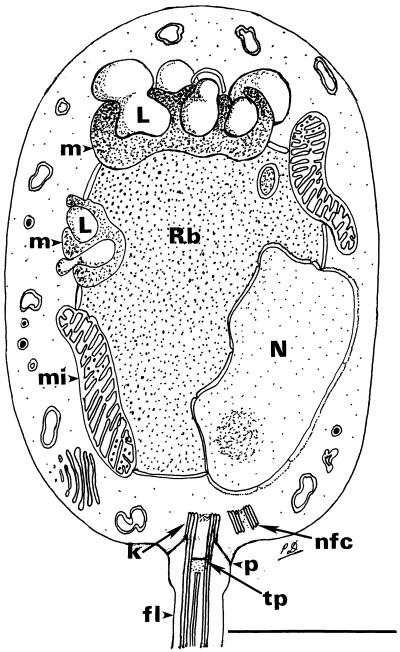
A composite line drawing of a longitudinal section through the zoospore of the chytridialean parasite found in the epidermis of Australian and Panamanian amphibians. The drawing is based on examination of thin sections of zoospores within the sporangium, not of serially sectioned, cultured, glutaraldehyde-fixed specimens. For these reasons, some details of the kinetosomal region were poorly preserved or may not have been visible. The unusual microbody–lipid globule complex and some details of the flagellar attachment are shown. In some sections, it appears that two microbody–lipid globule complexes are present, but it is unknown whether this represents a single complex wrapped around the ribosomal region. The flagellum in longitudinal and transverse section is drawn slightly larger than scale to adequately demonstrate the aspects of the flagellar attachment. Rb, ribosomal area; N, nucleus; L, lipid globule; m, microbody; mi, mitochondrion; k, kinetosome; p, prop attaching kinetosome to plasmalemma; tp, terminal plate; nfc, nonfunctioning centriole; fl, flagellum. (Bar = 1 μm.)

Maximum-likelihood tree inferred from ssu-rDNA sequences of the amphibian parasite and 44 other eukaryotes, based on 1,607 stably aligned positions at a transition/transversion ratio of 2.0. GenBank accession numbers of ssu-rDNAs other than the amphibian parasite in this tree have been published (23). Bootstrap values (percent of 1,000 replicates) are taken from the topologically identical parsimony tree. Distance matrices (corrected for superimposed substitutions by a generalized Kimura two-parameter model) were calculated by using dnadist from the phylip package, and neighbor-joining trees were inferred by using neighbor (24). Unweighted parsimony trees were inferred by using dnapars with 1,000 iterations (jumbles). Neighbor-joining and parsimony analyses were bootstrapped (n = 1000) by sequential use of seqboot , the inference program(s) above, and consense (24, 25). A maximum-likelihood tree was inferred by using dnaml . All inferences used random-addition order and, for parsimony and likelihood, global (43-level) optimization. The position of Apusomonas proboscidea ssu-rDNA was unstable, and in likelihood inference depended on the transition/transversion ratio.
References
-
- Blaustein A R, Wake D B. Trends Ecol Evol. 1990;5:203–204.
-
- Wake D B. Science. 1991;253:860. - PubMed
-
- Drost C A, Fellers G M. Conserv Biol. 1996;10:414–425.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
