

Control of cyclin B1 localization through regulated binding of the nuclear export factor CRM1
- PMID: 9679058
- PMCID: PMC317017
- DOI: 10.1101/gad.12.14.2131
Control of cyclin B1 localization through regulated binding of the nuclear export factor CRM1
Abstract
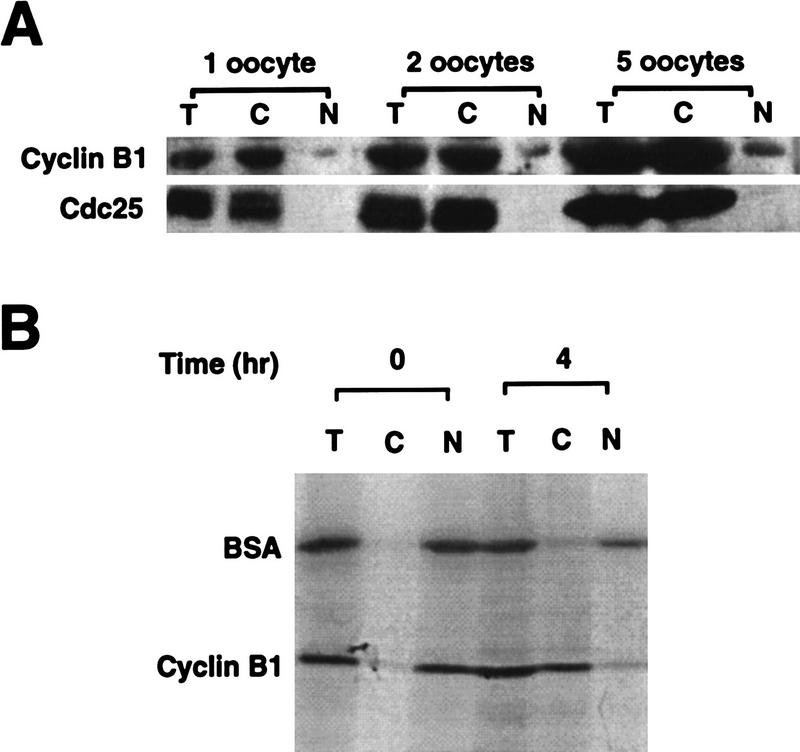
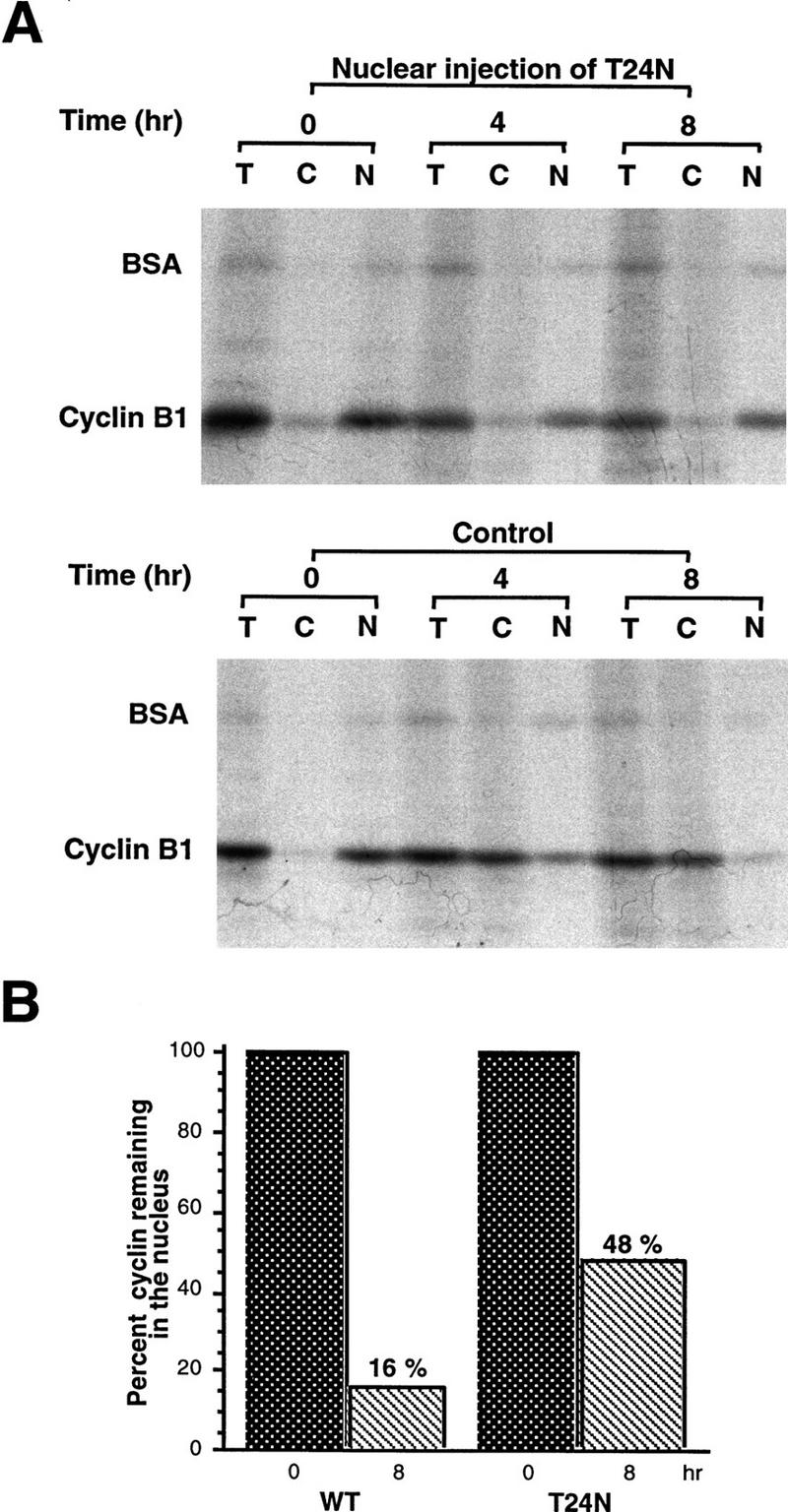
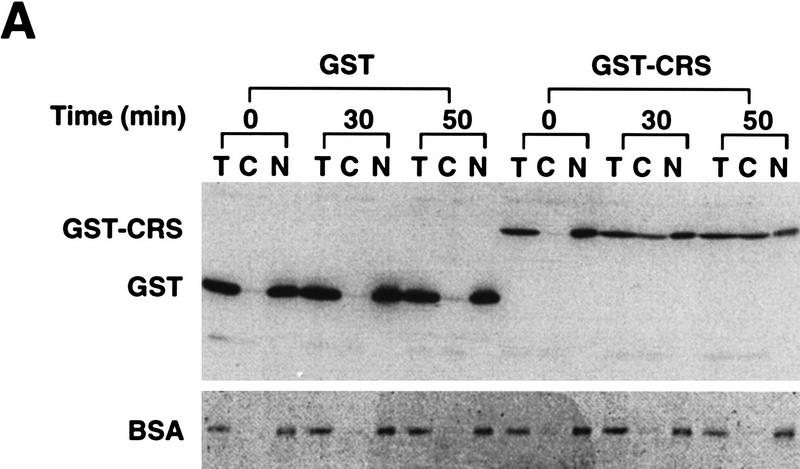
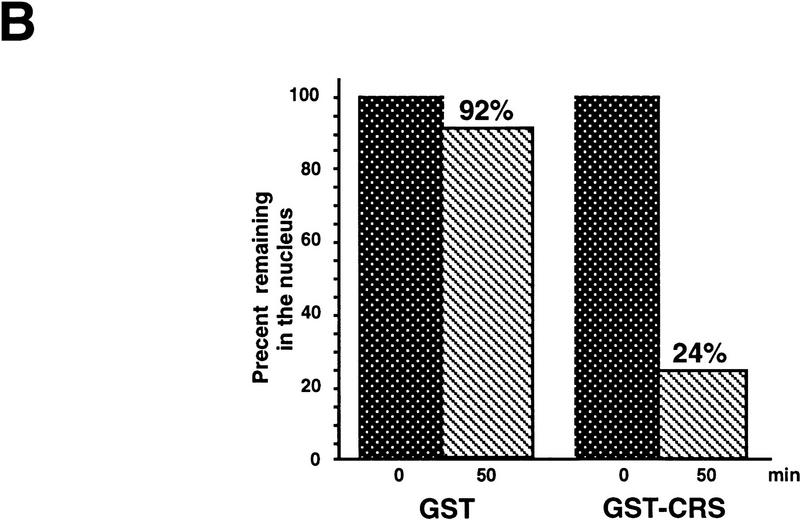
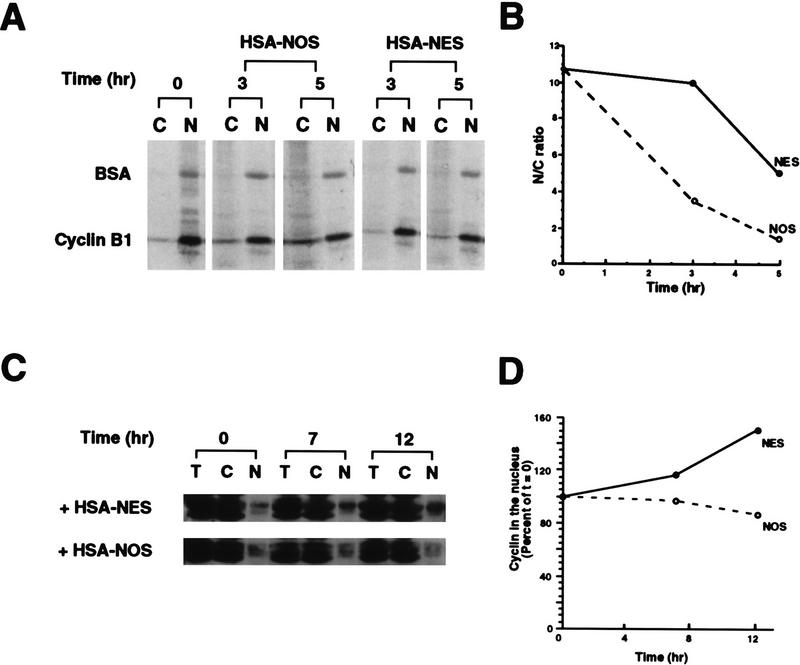
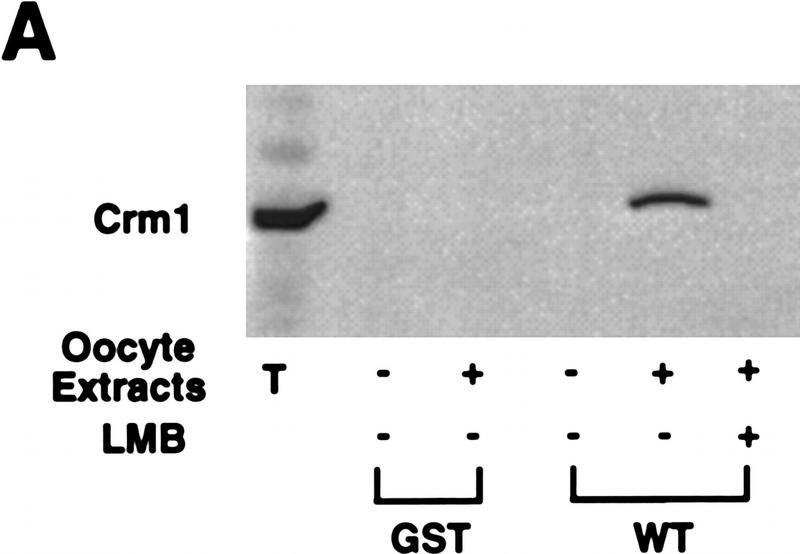
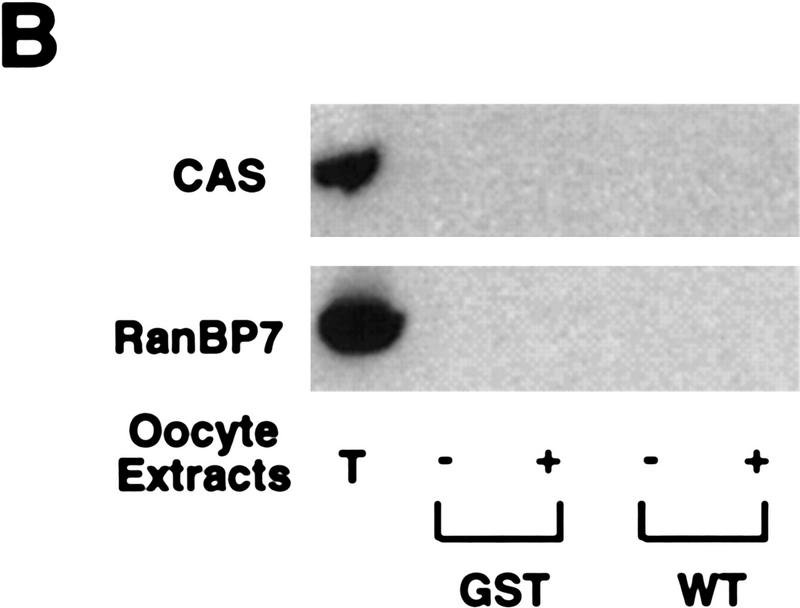
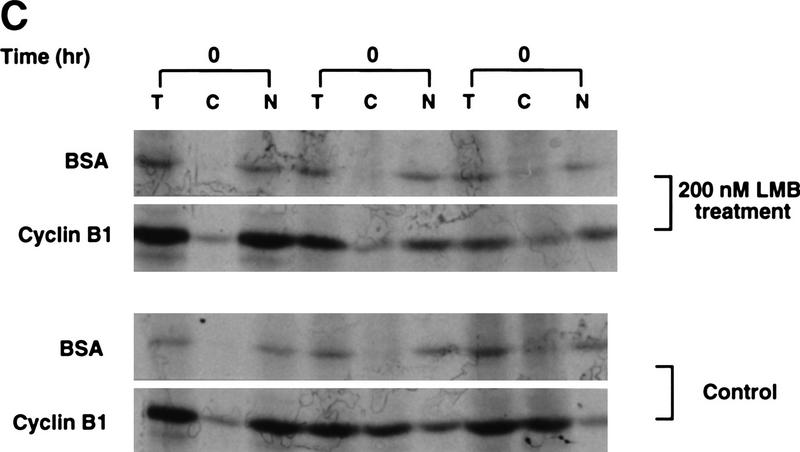
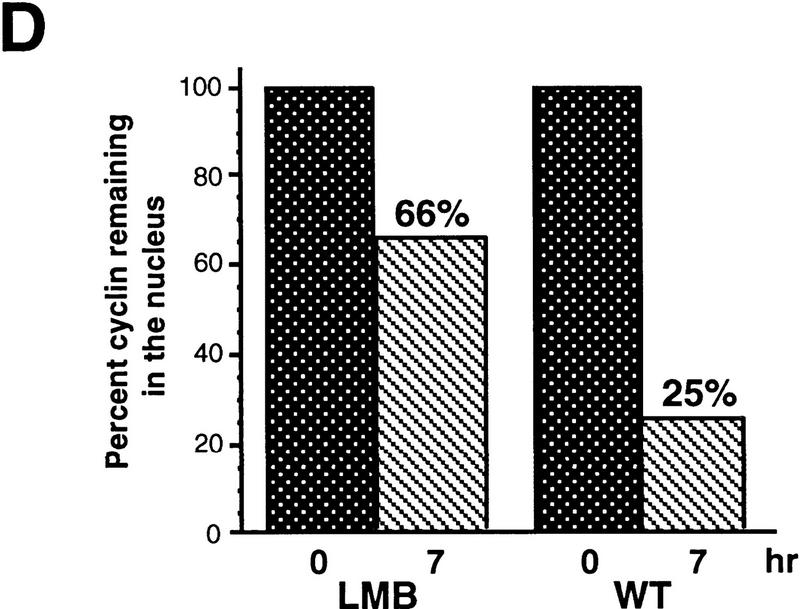
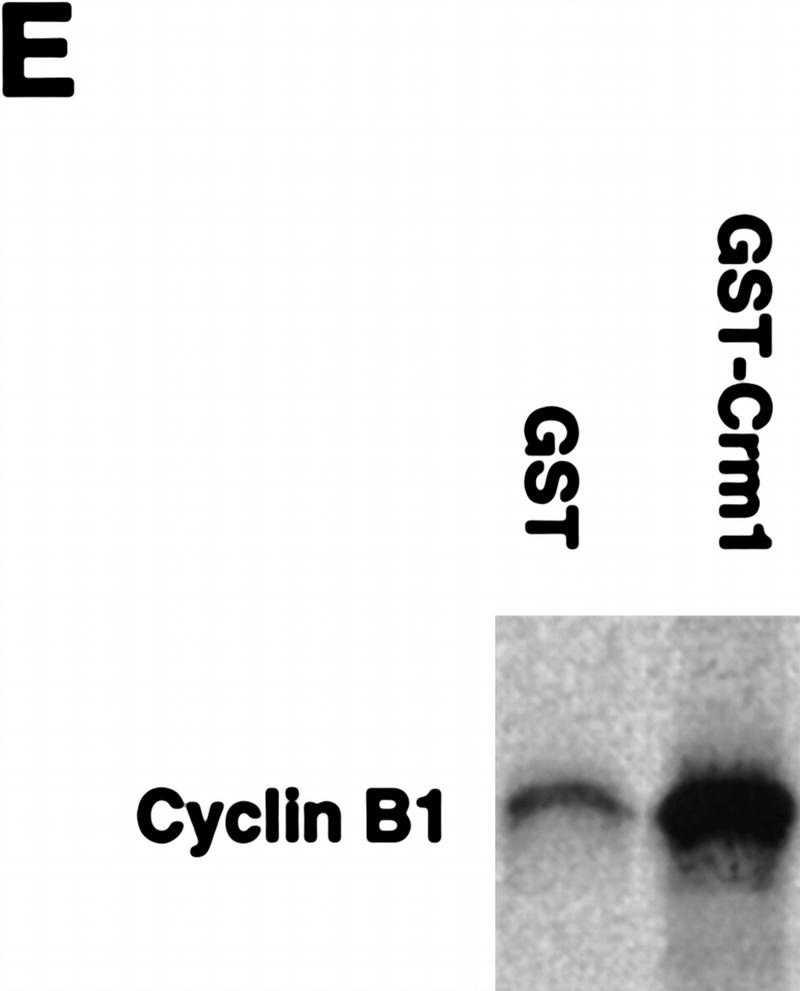
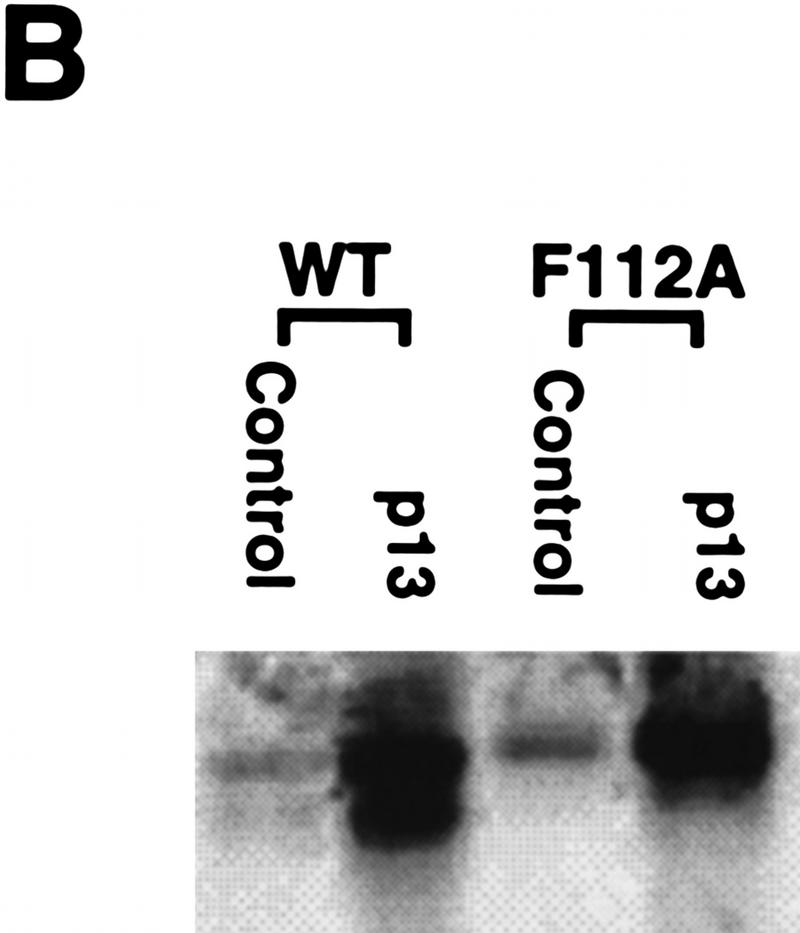
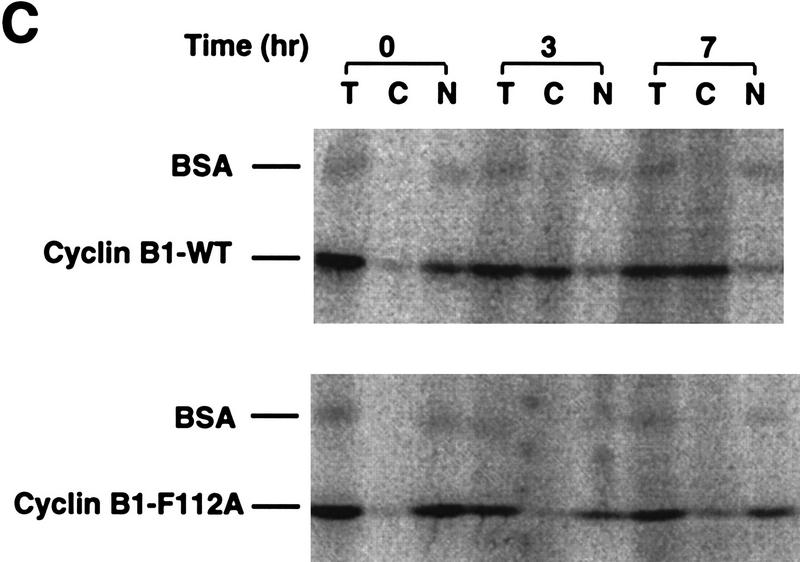
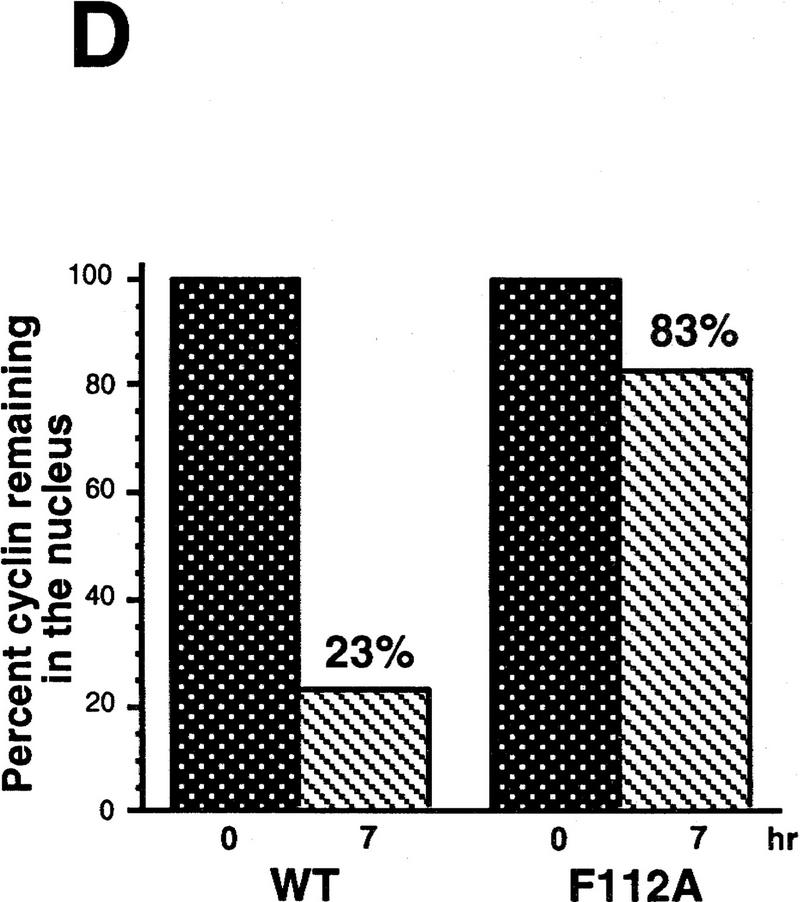
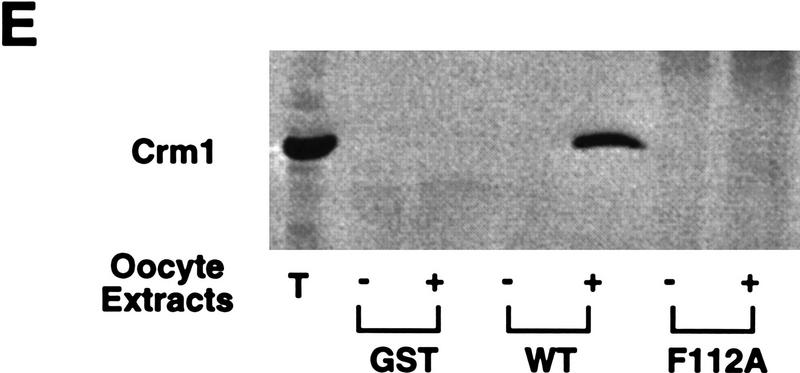
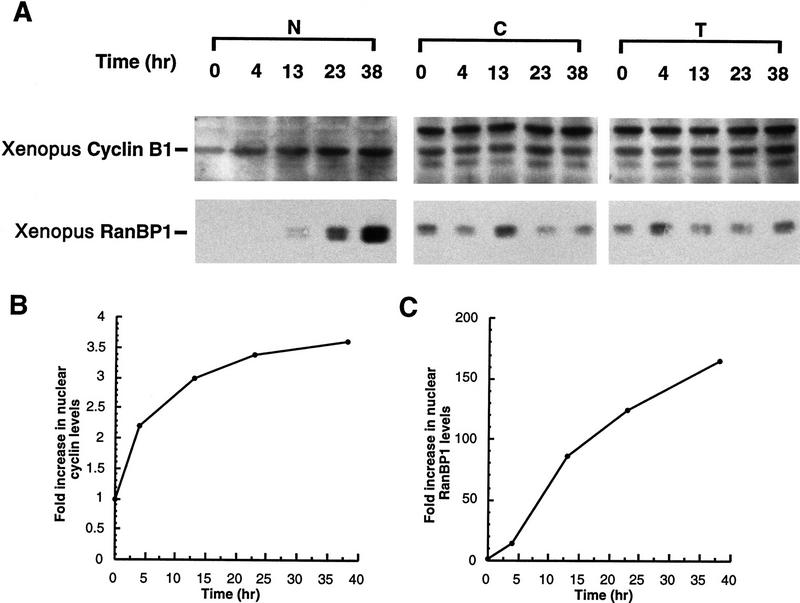
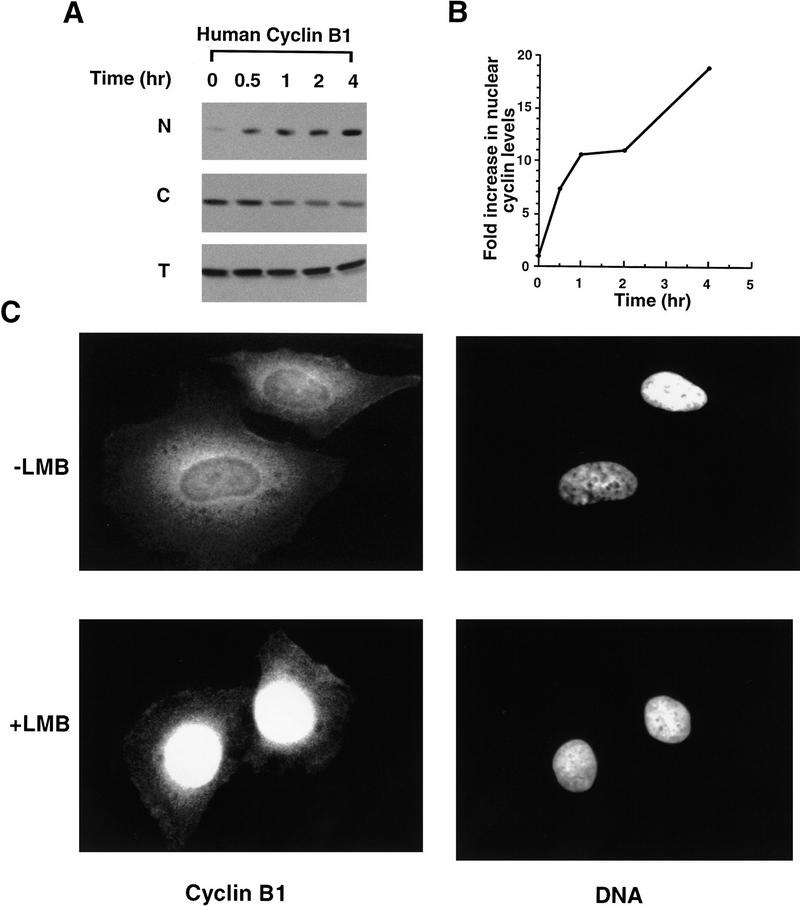
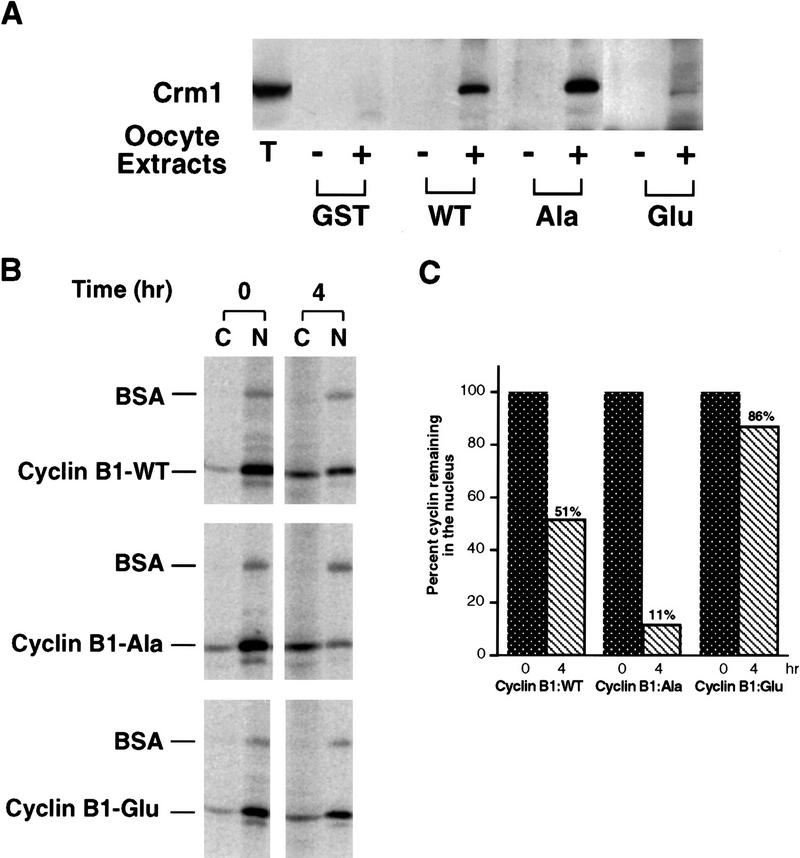
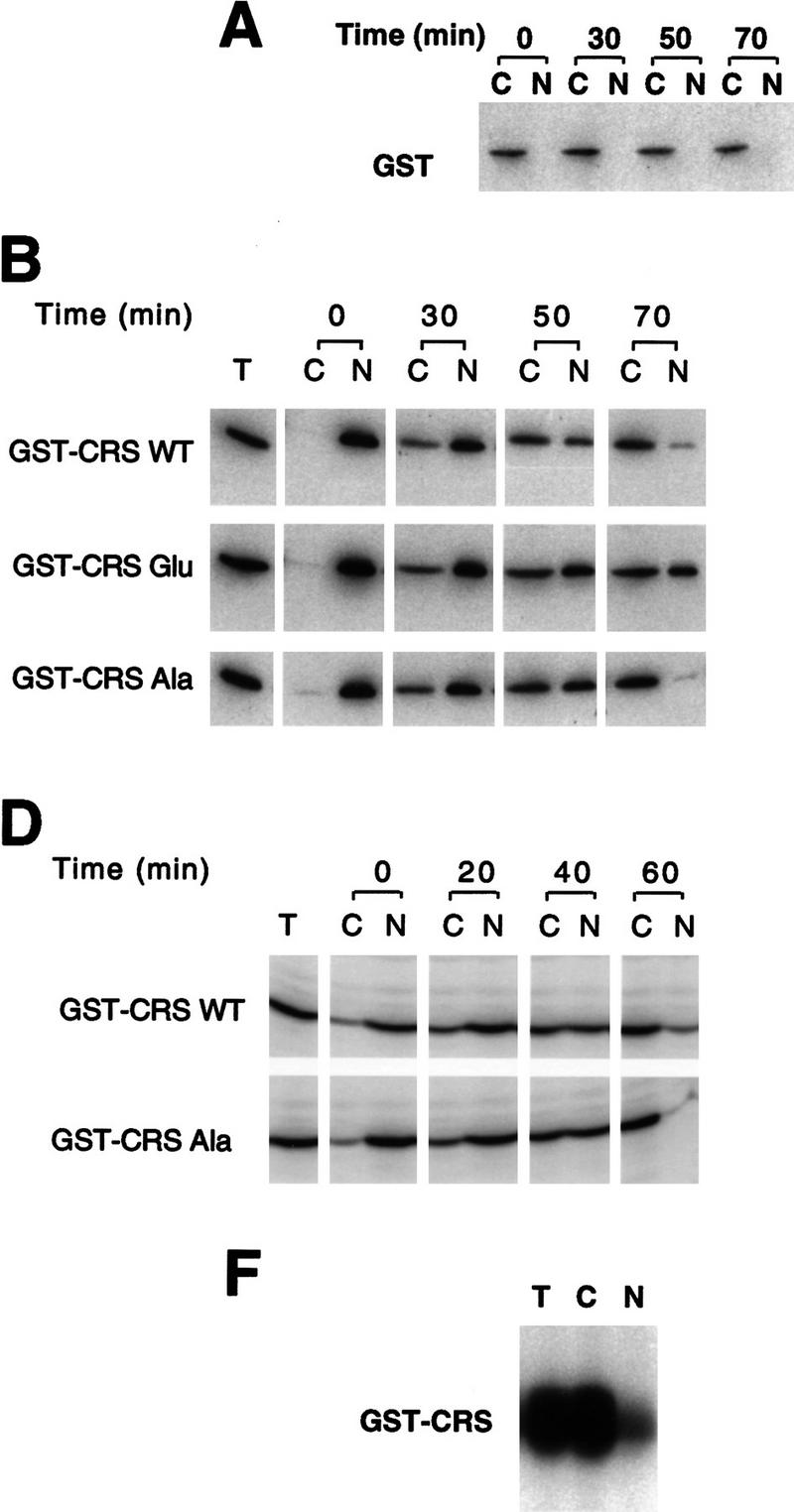
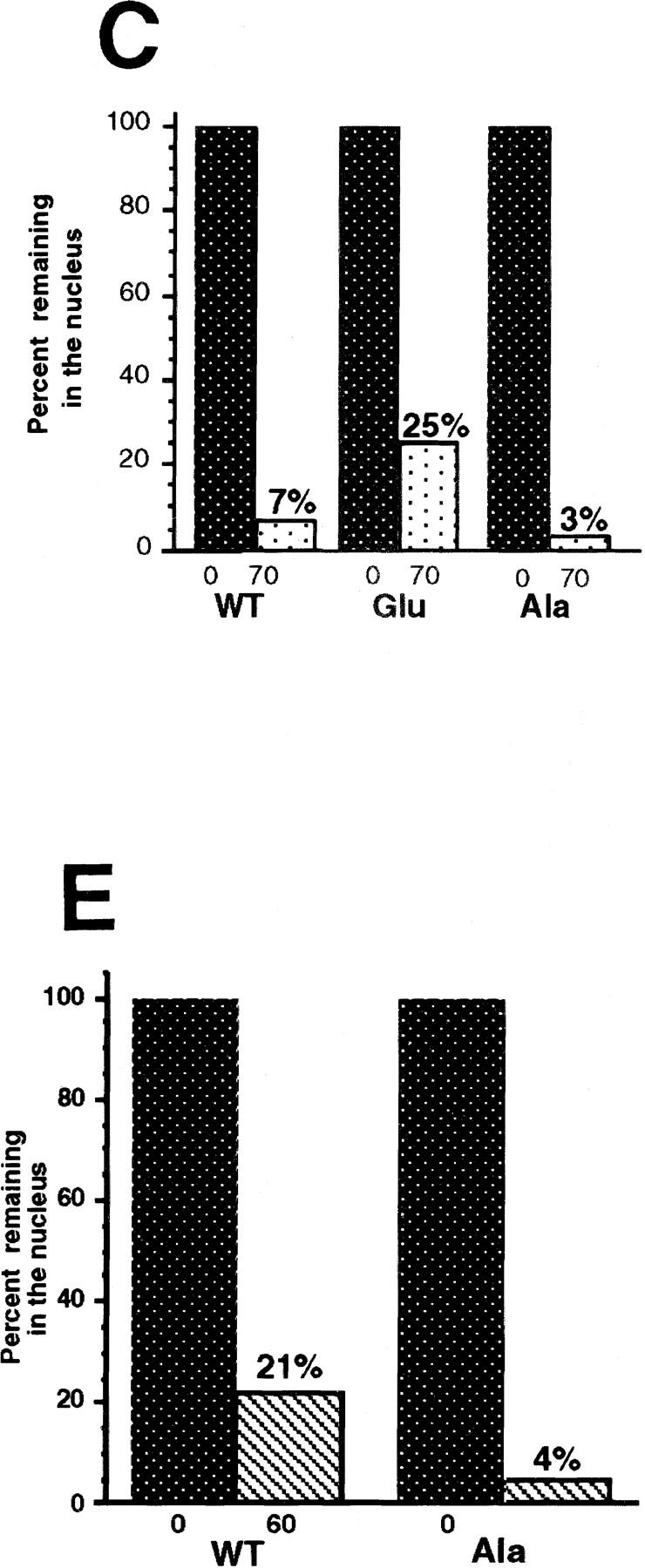
Activation of the Cyclin B/Cdc2 kinase complex triggers entry into mitosis in all eukaryotic cells. Cyclin B1 localization changes dramatically during the cell cycle, precipitously transiting from the cytoplasm to the nucleus at the beginning of mitosis. Presumably, this relocalization promotes the phosphorylation of nuclear targets critical for chromatin condensation and nuclear envelope breakdown. We show here that the previously characterized cytoplasmic retention sequence of Cyclin B1, responsible for its interphase cytoplasmic localization, is actually an autonomous nuclear export sequence, capable of directing nuclear export of a heterologous protein, and able to bind specifically to the recently identified export mediator, CRM1. We propose that the observed cytoplasmic localization of Cyclin B1 during interphase reflects the equilibrium between ongoing nuclear import and rapid CRM1-mediated export. In support of this hypothesis, we found that treatment of cells with leptomycin B, which disrupted Cyclin B1-CRM1 interactions, led to a marked nuclear accumulation of Cyclin B1. In mitosis, Cyclin B1 undergoes phosphorylation at several sites, a subset of which have been proposed to play a role in Cyclin B1 accumulation in the nucleus. Both CRM1 binding and the ability to direct nuclear export were affected by mutation of these phosphorylation sites; thus, we propose that Cyclin B1 phosphorylation at the G2/M transition prevents its interaction with CRM1, thereby reducing nuclear export and facilitating nuclear accumulation.
Figures

Similar articles
-
MPF localization is controlled by nuclear export.EMBO J. 1998 Jul 15;17(14):4127-38. doi: 10.1093/emboj/17.14.4127. EMBO J. 1998. PMID: 9670027 Free PMC article.
-
Combinatorial control of cyclin B1 nuclear trafficking through phosphorylation at multiple sites.J Biol Chem. 2001 Feb 2;276(5):3604-9. doi: 10.1074/jbc.M008151200. Epub 2000 Nov 1. J Biol Chem. 2001. PMID: 11060306
-
Nuclear export of cyclin B1 and its possible role in the DNA damage-induced G2 checkpoint.EMBO J. 1998 May 15;17(10):2728-35. doi: 10.1093/emboj/17.10.2728. EMBO J. 1998. PMID: 9582266 Free PMC article.
-
Cyclin B1 and CDK1: nuclear localization and upstream regulators.Prog Cell Cycle Res. 2003;5:335-47. Prog Cell Cycle Res. 2003. PMID: 14593728 Review.
-
Control of mitosis by changes in the subcellular location of cyclin-B1-Cdk1 and Cdc25C.Curr Opin Cell Biol. 2000 Dec;12(6):658-65. doi: 10.1016/s0955-0674(00)00149-6. Curr Opin Cell Biol. 2000. PMID: 11063929 Review.
Cited by
-
DNA damage checkpoint execution and the rules of its disengagement.Front Cell Dev Biol. 2022 Oct 6;10:1020643. doi: 10.3389/fcell.2022.1020643. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36274841 Free PMC article. Review.
-
Characterization of RanBPM molecular determinants that control its subcellular localization.PLoS One. 2015 Feb 6;10(2):e0117655. doi: 10.1371/journal.pone.0117655. eCollection 2015. PLoS One. 2015. PMID: 25659156 Free PMC article.
-
The biochemistry of mitosis.Cold Spring Harb Perspect Biol. 2015 Feb 6;7(3):a015776. doi: 10.1101/cshperspect.a015776. Cold Spring Harb Perspect Biol. 2015. PMID: 25663668 Free PMC article. Review.
-
Safeguarding entry into mitosis: the antephase checkpoint.Mol Cell Biol. 2010 Jan;30(1):22-32. doi: 10.1128/MCB.00687-09. Mol Cell Biol. 2010. PMID: 19841063 Free PMC article. Review.
-
An essential role for Ran GTPase in epithelial ovarian cancer cell survival.Mol Cancer. 2010 Oct 13;9:272. doi: 10.1186/1476-4598-9-272. Mol Cancer. 2010. PMID: 20942967 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous