

Functions of large conductance Ca2+-activated (BKCa), delayed rectifier (KV) and background K+ channels in the control of membrane potential in rabbit renal arcuate artery
- PMID: 9679171
- PMCID: PMC2231112
- DOI: 10.1111/j.1469-7793.1998.159bi.x
Functions of large conductance Ca2+-activated (BKCa), delayed rectifier (KV) and background K+ channels in the control of membrane potential in rabbit renal arcuate artery
Abstract
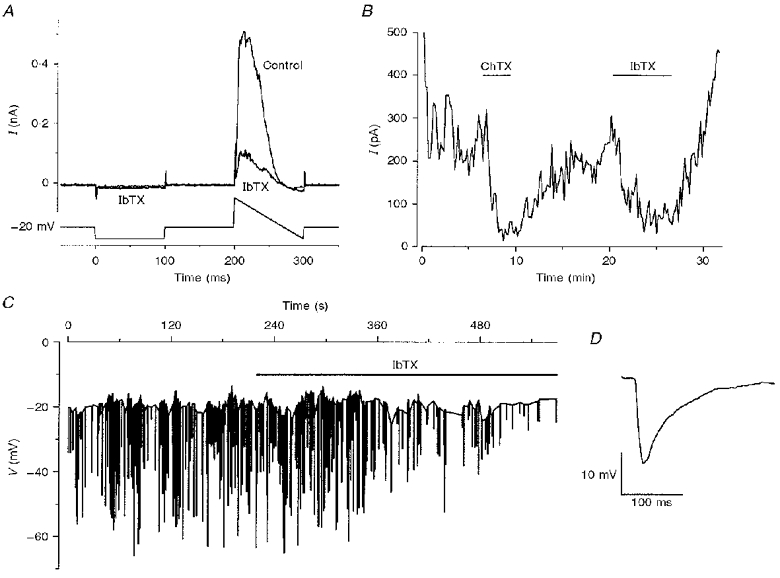
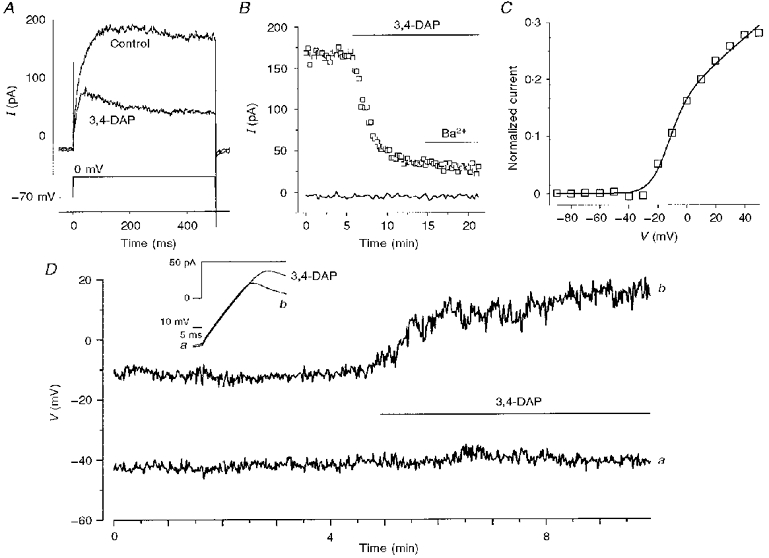
1. The types of K+ channel which determine the membrane potential of arcuate artery smooth muscle cells were investigated by patch-clamp recording from isolated cells and lumenal diameter measurements from intact pressurized renal arcuate arteries. 2. Single cells had a mean resting potential of -38 mV and were depolarized by 130 mM K+ but not by the Cl- channel blocker 4,4'-diisothiocyanatostilbene-2, 2'-disulphonic acid (DIDS). 3. Iberiotoxin did not affect the resting potential but inhibited spontaneous transient hyperpolarizations. Iberiotoxin or 1 mM tetraethylammonium (TEA+) constricted intact arteries. 3,4-Diaminopyridine (3,4-DAP)-sensitive delayed rectifier K+ (KV) channel current was elicited by depolarization but 3,4-DAP did not affect the resting potential or induce constriction in the intact artery. 4. A voltage-independent K+ current was inhibited by >= 0.1 mM barium (Ba2+) and unaffected by iberiotoxin, glibenclamide, apamin, 3,4-DAP and ouabain. In six out of ten cells, 1 mM Ba2+ depolarized the resting potential, while in the other cells the potential was resistant to all of the K+ channel blockers and ouabain. Ba2+ (0.1-1 mM) constricted the intact artery, but 10 microM Ba2+, 1 microM glibenclamide or 100 nM apamin had no effect. 5. The data suggest that resting potential is determined by background K+ channels, one type being Ba2+ sensitive and voltage independent, and another type being poorly defined due to its resistance to any inhibitor. Large conductance Ca2+-activated K+ (BKCa) and KV channels do not determine the resting potential but have separate functions to underlie transient Ca2+-induced hyperpolarizations and to protect against depolarization past about -30 mV.
Figures




Similar articles
-
Role of K+ channels in A2A adenosine receptor-mediated dilation of the pressurized renal arcuate artery.Br J Pharmacol. 1999 Jan;126(2):494-500. doi: 10.1038/sj.bjp.0702310. Br J Pharmacol. 1999. PMID: 10077243 Free PMC article.
-
Effects of the BKCa channel activator, NS1619, on rat cerebral artery smooth muscle.Br J Pharmacol. 1996 Jan;117(1):119-29. doi: 10.1111/j.1476-5381.1996.tb15163.x. Br J Pharmacol. 1996. PMID: 8825352 Free PMC article.
-
A comparison of EDHF-mediated and anandamide-induced relaxations in the rat isolated mesenteric artery.Br J Pharmacol. 1997 Dec;122(8):1573-84. doi: 10.1038/sj.bjp.0701546. Br J Pharmacol. 1997. PMID: 9422801 Free PMC article.
-
Changes in the expression and function of arterial potassium channels during hypertension.Vascul Pharmacol. 2002 Jan;38(1):13-23. doi: 10.1016/s1537-1891(02)00122-2. Vascul Pharmacol. 2002. PMID: 12378818 Review.
-
Physiological roles of K+ channels in vascular smooth muscle cells.J Smooth Muscle Res. 2008 Apr;44(2):65-81. doi: 10.1540/jsmr.44.65. J Smooth Muscle Res. 2008. PMID: 18552454 Review.
Cited by
-
Role of K+ channels in A2A adenosine receptor-mediated dilation of the pressurized renal arcuate artery.Br J Pharmacol. 1999 Jan;126(2):494-500. doi: 10.1038/sj.bjp.0702310. Br J Pharmacol. 1999. PMID: 10077243 Free PMC article.
-
Expression and function of native potassium channel [K(V)alpha1] subunits in terminal arterioles of rabbit.J Physiol. 2001 Aug 1;534(Pt 3):691-700. doi: 10.1111/j.1469-7793.2001.00691.x. J Physiol. 2001. PMID: 11483700 Free PMC article.
-
Complex interactions of NO/cGMP/PKG systems on Ca2+ signaling in afferent arteriolar vascular smooth muscle.Am J Physiol Heart Circ Physiol. 2010 Jan;298(1):H144-51. doi: 10.1152/ajpheart.00485.2009. Epub 2009 Oct 30. Am J Physiol Heart Circ Physiol. 2010. PMID: 19880669 Free PMC article.
-
Two-pore potassium channels in the cardiovascular system.Eur Biophys J. 2009 Mar;38(3):305-18. doi: 10.1007/s00249-008-0326-8. Epub 2008 May 1. Eur Biophys J. 2009. PMID: 18449536 Review.
-
Closure of multiple types of K+ channels is necessary to induce changes in renal vascular resistance in vivo in rats.Pflugers Arch. 2011 Nov;462(5):655-67. doi: 10.1007/s00424-011-1018-2. Epub 2011 Aug 27. Pflugers Arch. 2011. PMID: 21874333
References
-
- Archer SL, Huang JMC, Reeve HL, Hampl V, Tolarová S, Michelakis E, Weir EK. Differential distribution of electrophysiologically distinct myocytes in conduit and resistance arteries determines their response to nitric oxide and hypoxia. Circulation Research. 1996;78:431–442. - PubMed
-
- Bolton TB, Imaizumi Y. Spontaneous transient outward currents in smooth muscle cells. Cell Calcium. 1996;20:141–152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous