

Lyn, Jak2, and Raf-1 kinases are critical for the antiapoptotic effect of interleukin 5, whereas only Raf-1 kinase is essential for eosinophil activation and degranulation
- PMID: 9687520
- PMCID: PMC2212466
- DOI: 10.1084/jem.188.3.421
Lyn, Jak2, and Raf-1 kinases are critical for the antiapoptotic effect of interleukin 5, whereas only Raf-1 kinase is essential for eosinophil activation and degranulation
Abstract
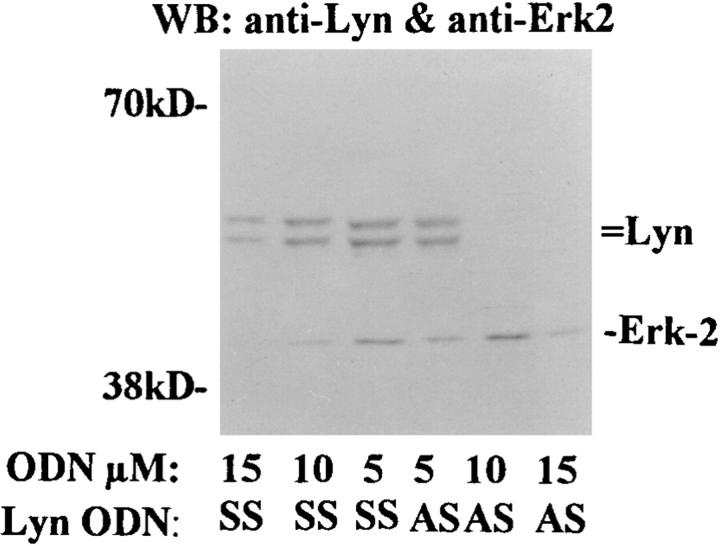
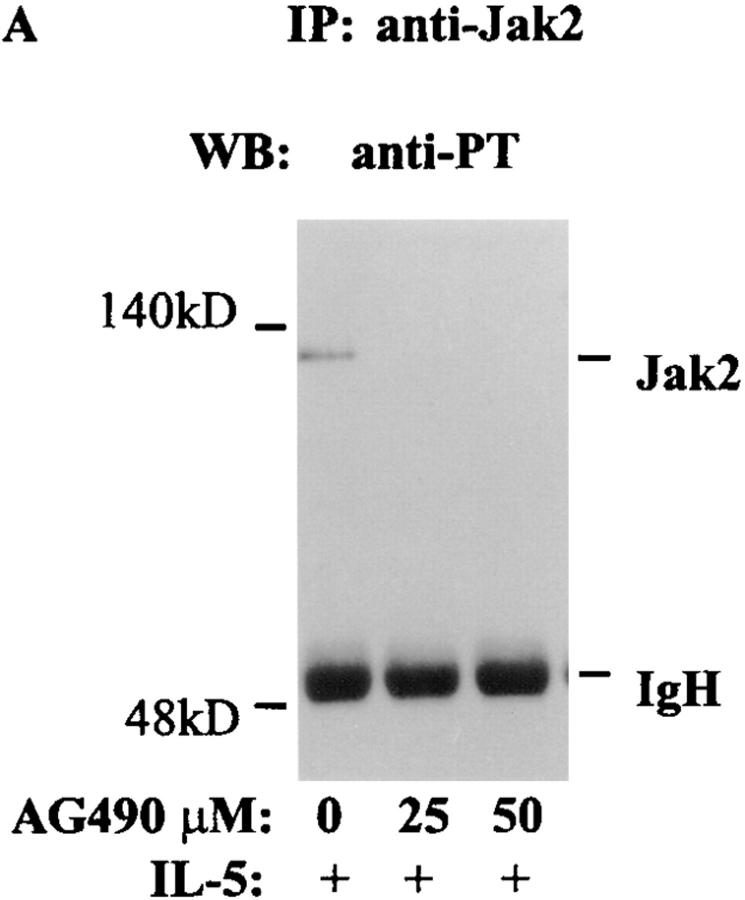
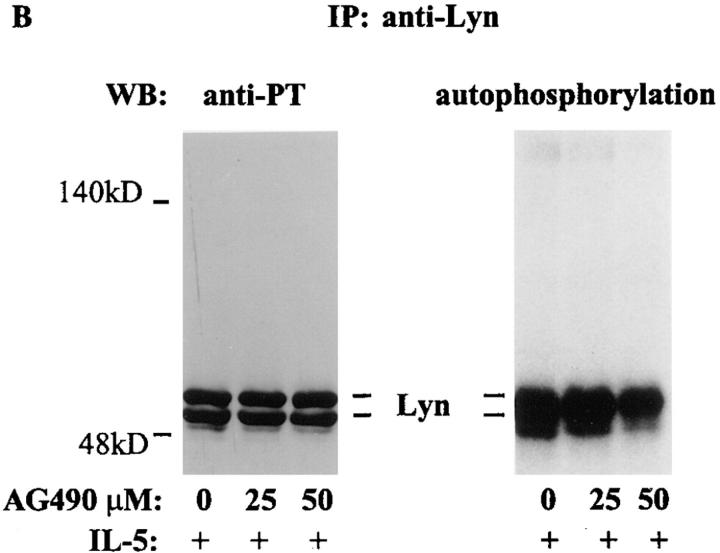
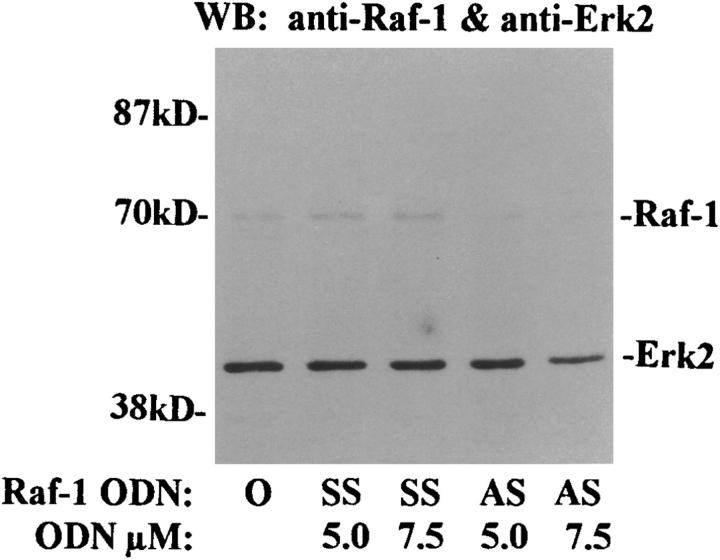
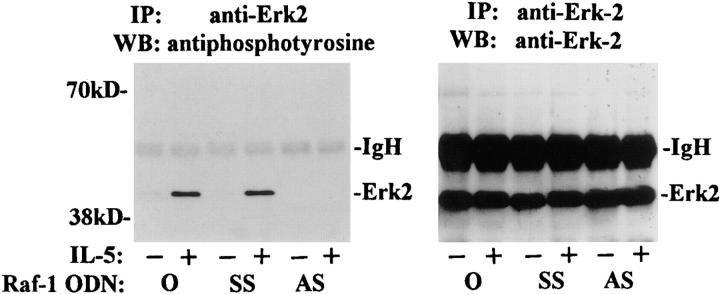
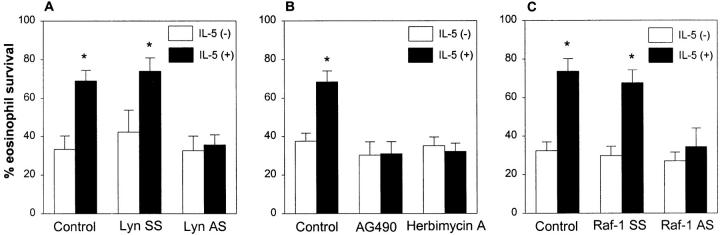
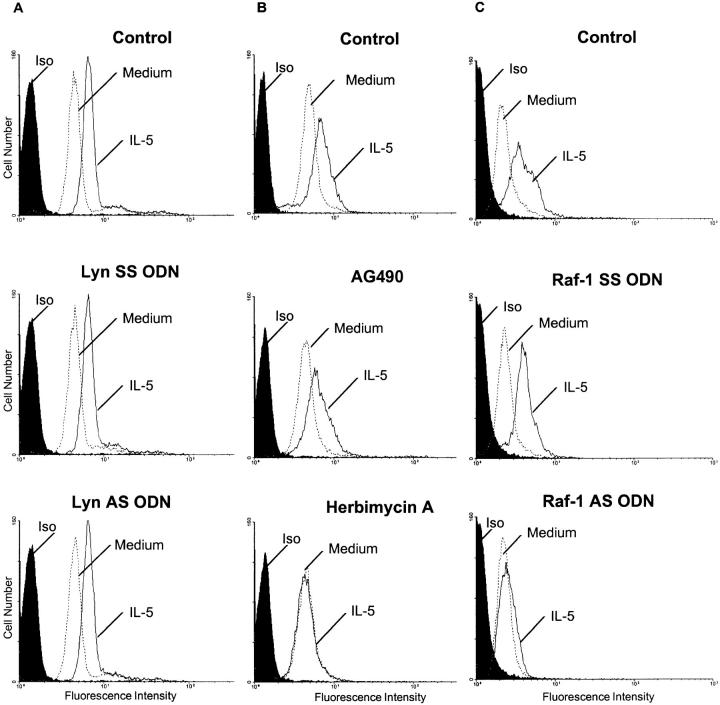
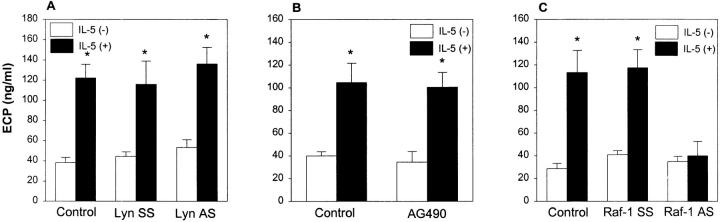
Interleukin (IL)-5 has been shown to activate many signaling molecules in eosinophils, but their functional relevance remains unknown. We have examined the functional relevance of Lyn, Jak2, and Raf-1 kinases in eosinophil survival, upregulation of adhesion molecules and degranulation. To this goal we used Lyn and Raf-1 antisense (AS) oligodeoxynucleotides (ODN) to inhibit the expression of these proteins and tyrphostin AG490 to specifically block the activation of Jak2. We have demonstrated that all three kinases are important for IL-5- induced suppression of eosinophil apoptosis. However, Lyn and Jak2 tyrosine kinases are not important for the upregulation of CD11b and the secretion of eosinophil cationic protein. In contrast, Raf-1 kinase is critical for both these functions. This is the first identification of specific signaling molecules responsible for three important functions of eosinophils. We have established a central role for Raf-1 kinase in regulating eosinophil survival, expression of beta2 integrins and degranulation. Further, there appears to be a dissociation between two receptor-associated tyrosine kinases, i.e., Lyn and Jak2, and the activation of Raf-1 kinase. The delineation of the functional relevance of signaling molecules will help design therapeutic approaches targeting specific eosinophil function.
Figures
References
-
- Corrigan CJ, Kay AB. T cells and eosinophils in the pathogenesis of asthma. Immunol Today. 1992;13:501–503. - PubMed
-
- Owen, W.F., Jr., and K.F. Austen. 1994. Cytokine regulation of eosinophil-mediated inflammatory reactions by modulation of eosinophil programmed cell death and subsequent priming for augmented function. In Eosinophils In Allergy and Inflammation. G.J. Gleich and A.B. Kay, editors. Marcel Decker, New York. 239–253.
-
- Resnick MB, Weller PF. Mechanisms of eosinophil recruitment. Am J Res Cell Mol Biol. 1993;8:349–355. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
