

Detection of subtle phenotypes: the case of the cell adhesion molecule csA in Dictyostelium
- PMID: 9689085
- PMCID: PMC21343
- DOI: 10.1073/pnas.95.16.9360
Detection of subtle phenotypes: the case of the cell adhesion molecule csA in Dictyostelium
Abstract
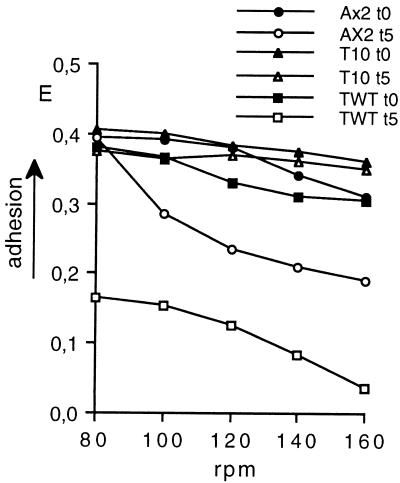
Dictyostelium amoebae aggregate into a multicellular organism by cAMP-driven chemotaxis and cell-cell adhesion. Cell adhesion is mediated by an EDTA-sensitive and an EDTA-resistant adhesion system. The latter is developmentally regulated and triggered by homophilic interactions of the membrane glycoprotein csA; on disruption of the encoding gene, EDTA-resistant contacts fail to form. Nevertheless, csA-null cells under usual laboratory conditions aggregate normally and complete development. By using experimental conditions that reproduce more closely the habitat of Dictyostelium amoebae, evidence is provided that csA is required for development and that its expression confers a selective advantage to populations of wild-type cells over csA-null mutants. The latter display reduced cell-cell adhesion, increased adhesiveness to the substratum, and slower motility, which lead to their sorting out from aggregating wild-type cells. It is proposed that the experimental conditions commonly used in the laboratory are not stringent enough to assess the developmental role of csA and other proteins. The assay described can be used to detect subtle phenotypes, to reexamine the developmental role of apparently nonessential genes, and to test the validity of recent models on emergence and maintenance of apparent genetic redundancy.
Figures
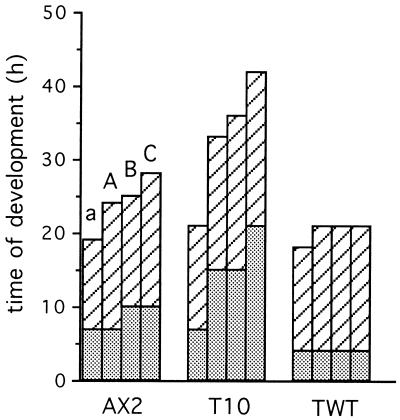
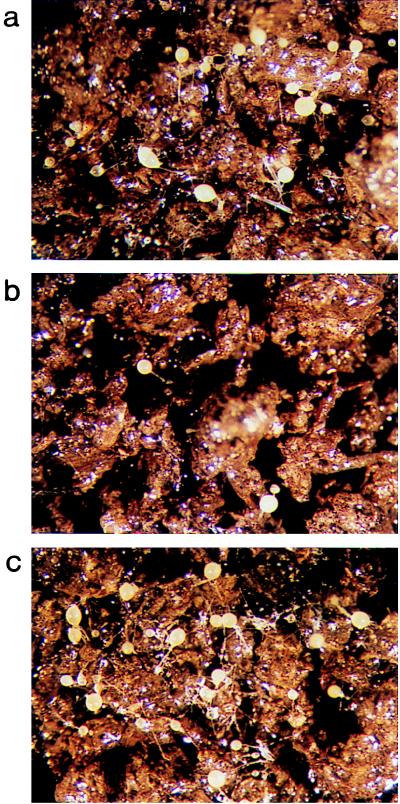
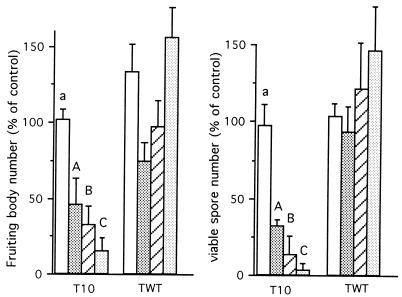
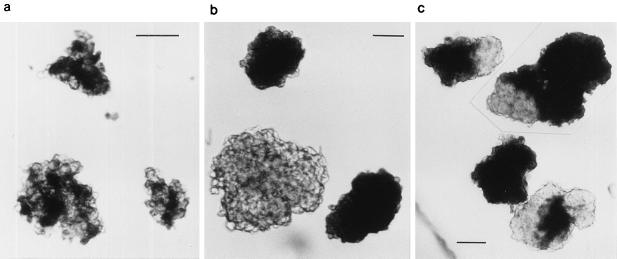
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
