

Trichostatin A causes selective loss of DNA methylation in Neurospora
- PMID: 9689097
- PMCID: PMC21355
- DOI: 10.1073/pnas.95.16.9430
Trichostatin A causes selective loss of DNA methylation in Neurospora
Abstract
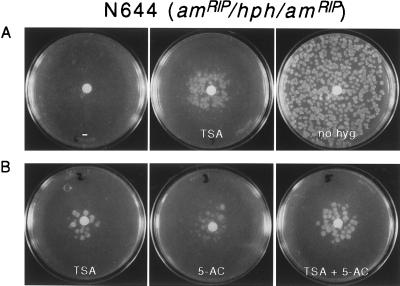
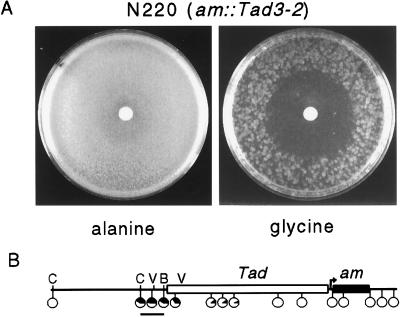
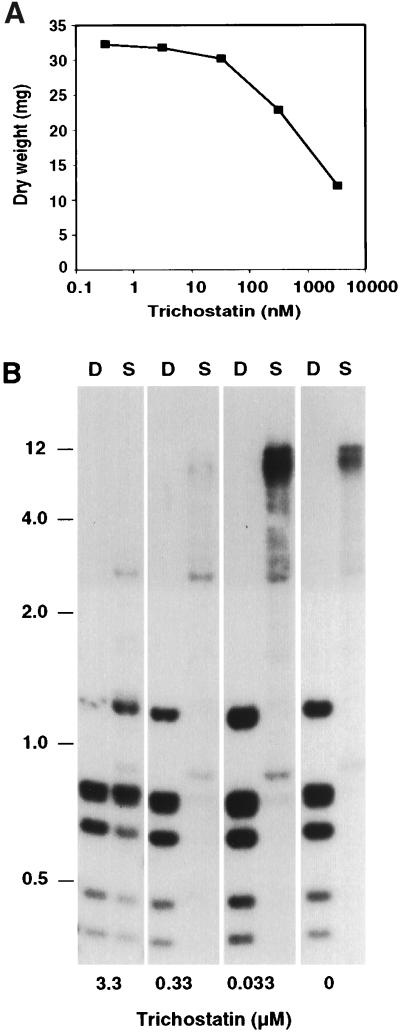
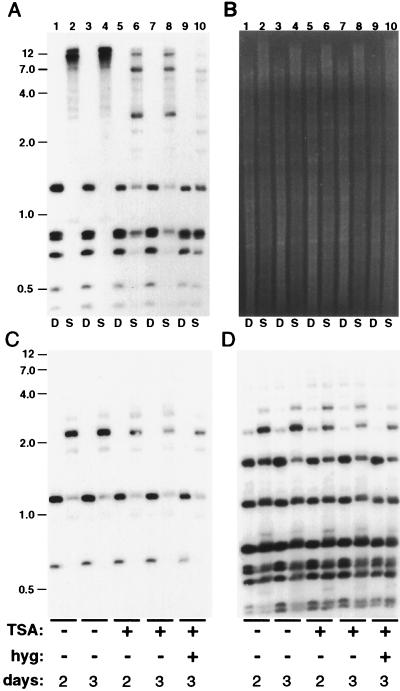
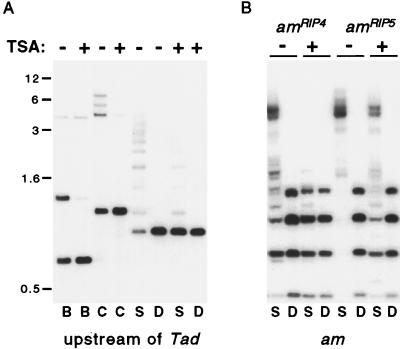
Both DNA methylation and hypoacetylation of core histones are frequently associated with repression of gene expression. Possible connections between these processes were investigated by taking advantage of genes controlled by methylation in Neurospora crassa. Trichostatin A (TSA), a potent inhibitor of histone deacetylase, derepressed a copy of hph that was repressed by DNA methylation which resulted from repeat-induced point mutation (RIP) acting on sequences flanking hph. Derepression by TSA was comparable to derepression by the inhibitor of DNA methylation, 5-azacytidine. TSA treatment also repressed an allele of am whose expression depends on methylation of an adjacent transposon, Tad. DNA methylation in the hph and Tad/am regions was greatly reduced by TSA treatment. TSA also caused hypomethylation of other methylated alleles of am generated by RIP. In contrast, TSA did not affect methylation of several other methylated genomic sequences examined, including the nucleolar rDNA and the inactivated transposon PuntRIP1. Several possible models are discussed for the observed selective demethylation induced by TSA. The implication that acetylation of chromatin proteins can directly or indirectly control DNA methylation raises the possibility that connections between protein acetylation and DNA methylation result in self-reinforcing epigenetic states.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
