

doi: 10.1159/000017314.
Molecular components of striatal plasticity: the various routes of cyclic AMP pathways
Affiliations
- PMID: 9691194
- PMCID: PMC4205584
- DOI: 10.1159/000017314
Item in Clipboard
Molecular components of striatal plasticity: the various routes of cyclic AMP pathways
Dev Neurosci.
1998.
Abstract
Neuroplasticity serves an important role for normal striatal function and in disease states. One route to neuroplasticity involves activation of the transcription factor cyclic 3', 5'-adenosine monophosphate (cyclic AMP) response element binding protein (CREB) by phosphorylation of the amino acid 133Ser. Dopamine and glutamate, the two predominant neurotransmitters in the striatum, induce CREB phosphorylation in primary cultures of rat striatum through cyclic AMP and Ca2+ pathways. Here we present the role of N-methyl-D-aspartate receptors and Ca2+ in cyclic AMP-mediated CREB phosphorylation.
Figures
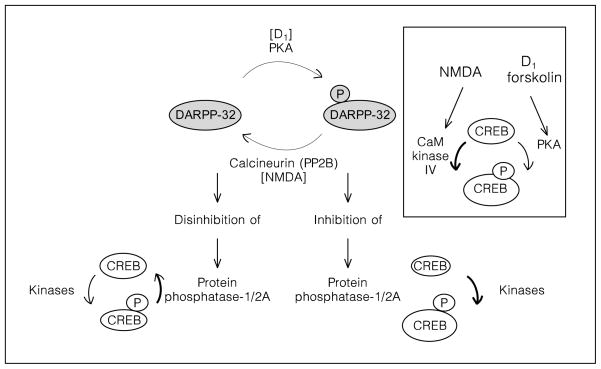
Role of dopamine/cyclic AMP and NMDA/Ca2+ in the activation of protein phosphatases and dephosphorylation of 133Ser-CREB in the striatum. The level of CREB phosphorylation is determined by the relative activity of kinases and phosphatases. The phosphatases responsible for dephosphorylation of 133Ser-CREB are PP1 and PP2A [65, 66]. The DARPP-32 pathway is involved in the regulation of PP1/2A in the striatum [55]. DARPP-32 is a neuronal phosphoprotein which upon phosphorylation on 34Thr [54, 111, 112], becomes a very potent PP1/2A inhibitor [55]. Dopamine-activated PKA phosphorylates DARPP-32 on 34Thr. Therefore, dopamine and cyclic AMP activation promotes CREB phosphorylation via inhibition of phosphatase activity. Stimulation of NMDA receptors induces the calcium-dependent phosphatase calcineurin, the dephosphorylation of DARPP-32 on 34Thr, the disinhibition of PP1/2A [52, 56, 111] and the dephosphorylation of CREB [66]. Insert: CaM kinase IV activated by NMDA receptors [75, 113], and PKA activated by D1 receptors [30, 57, 60, 114] are involved in the phosphorylation of CREB on 133Ser.
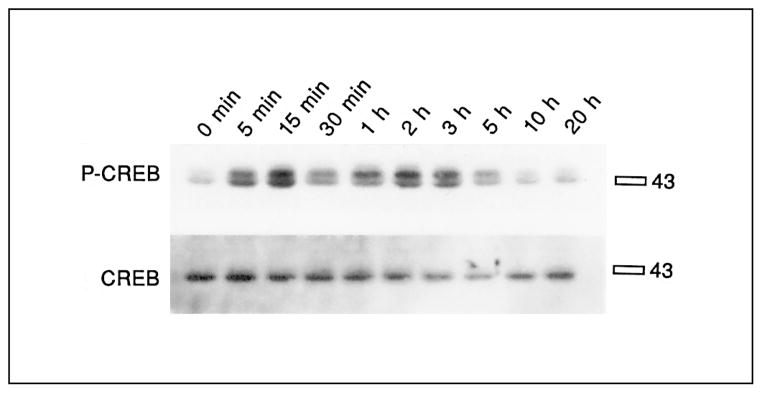
Time course of CREB phosphorylation after stimulation with forskolin in primary striatal cultures. Primary striatal cultures were treated with forskolin (20 μM) for the times indicated, medium was removed and cells were frozen in liquid nitrogen until the day of the experiment. The immunoblot was developed with 133Ser-phos-pho-CREB antibody (upper blot), and reprobed with CREB antiserum (lower blot). The specific bands ran slightly above the 43-kD marker. No significant difference of 133Ser-phospho-CREB induction was observed between 5 min and 3 h. No significant differences of CREB levels were observed at any time points (n = 2).
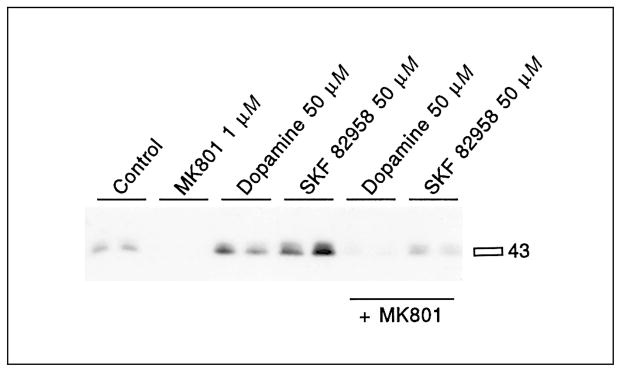
Dopamine and SKF82958-mediated 133Ser-CREB phosphorylation is inhibited by the NMDA antagonist MK801. MK801 (1 μM) was added 30 min before dopamine (50 μM) or SKF82958 (50 μM). Both dopamine and SKF82958 induced 133Ser-CREB phosphorylation. Pretreatment with MK801 for 30 min prevented the induction of CREB phosphorylation. See table 1 for statistical analyses.
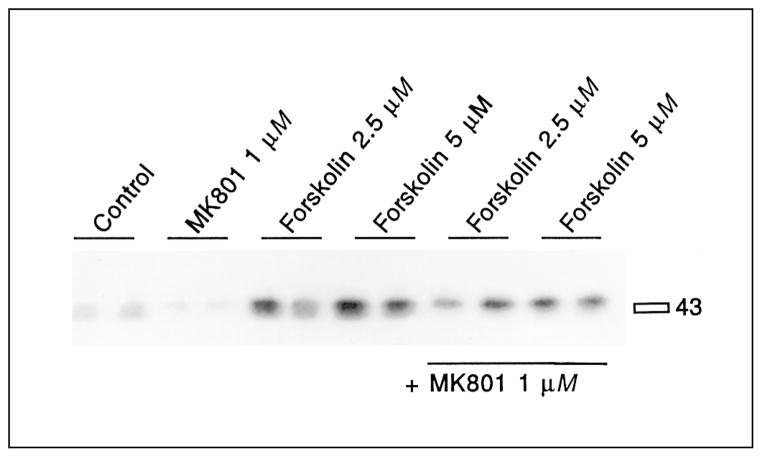
CREB phosphorylation mediated by low concentrations of forskolin is inhibited by the NMDA antagonist MK801. Cells exposed to 2.5 or 5 μM forskolin show increased CREB phosphorylation. This increase is blocked by pretreatment with MK801 (1 μM) for 30 min (last 4 lanes). See table 2 for statistical analyses.
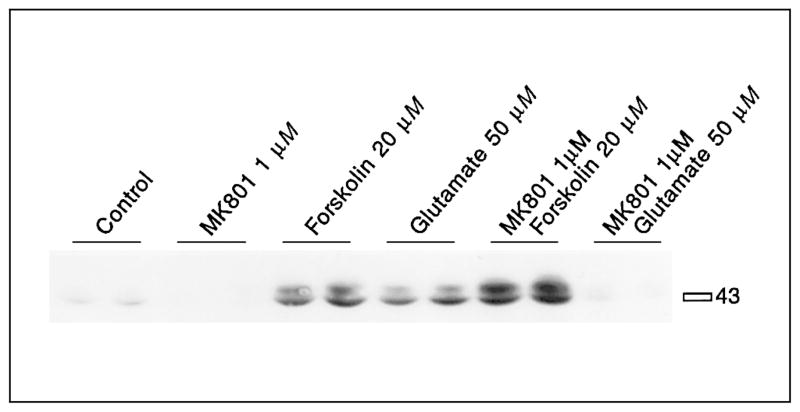
CREB phosphorylation mediated by high concentrations of forskolin is not inhibited by the NMDA antagonist MK801. Cells exposed to 20 μM forskolin show increased CREB phosphorylation, which is further increased after MK801 (1 μM) treatment. Glutamate-mediated CREB phosphorylation is blocked by MK801. MK801 was added 30 min before forskolin (20 μM). See table 3 for statistical analyses.
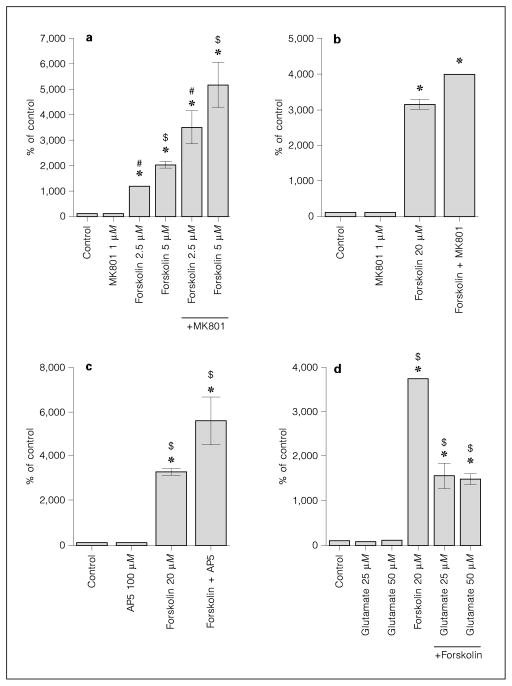
NMDA antagonists increase forskolin-activated cyclic AMP levels. a Forskolin 2.5 μM and forskolin 5 μM increase cyclic AMP levels in a dose-dependent manner. Pretreatment with MK801 (1 μM) augments forskolin-induced cyclic AMP levels. Analysis done in quadruplicates. b Forskolin 20 μM-induced cyclic AMP levels are augmented by MK801 (1 μM). Analysis done in pentuplicates. c Forskolin (20 μM)-induced cyclic AMP levels are augmented by pre-treatment with AP5 (100 μM). Analysis done in quadruplicates. d Glutamate (25 and 50 μM) decreases cyclic AMP levels induced by forskolin (20 μM). Analysis done in quadruplicates. MK801, AP5 and glutamate were added 30 min before forskolin. All data presented as average ± SD in % of untreated control. Some error bars are below the resolution of the graphs. * = Significantly different from control (Dunnett’s method); #, $ = significant difference between groups (Tukey-Kramer HSD).
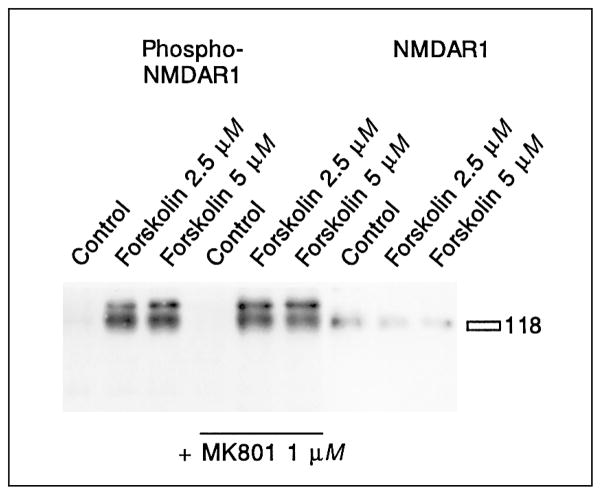
Modulation of NMDA receptor phosphorylation by forskolin. Cells treated with forskolin show phosphorylation of the NMDAR1 protein, as revealed in an immunoblot with a phospho-NMDAR1 receptor antibody. MK801 does not block phosphorylation. All nine samples were run in one gel, transferred, and the membrane was cut before the last three lanes. The left side was developed with an antibody specific for the phosphorylated form of the NMDAR1 receptor, the right side was developed with an antibody specific for the NMDAR1 receptor. The NMDAR1 receptor runs slightly above the 118-kD marker. The phospho-NMDAR1 receptor antibody stains 2 bands around 120 kD, the smaller of which is also detected by the NMDAR1 antiserum and was considered the specific band. See table 4 for statistical analyses.
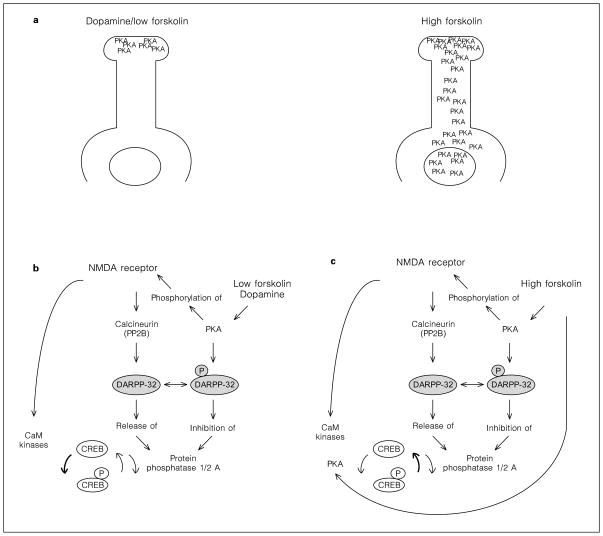
Cyclic AMP mediates CREB phosohorylation by two different mechanism. a The distribution of activated PKA after stimulation with low levels of forskolin or dopamine, and after high levels of forskolin. b Low levels of forskolin increase local PKA activity which causes phosphorylation of the NMDAR1 receptor. Kinases activated intraneuronally by the NMDA receptor pathway phosphorylate CREB. NMDA receptors also stimulate PP1 activity through the DARPP-32 pathway, but the equilibrium favors kinase activity, as demonstrated by increased 133Ser-CREB phosphorylation. Therefore, inhibition of NMDA receptors prevents CREB phosphorylation. The bold arrow points out the predominant action of the NMDA receptor pathway in the presence of low concentrations of forskolin. c High levels of forskolin stimulate sufficient amounts of PKA to directly phosphorylate 133Ser-CREB. Kinases activated by NMDA receptors cannot further contribute to CREB phosphorylation, while the activation of calcineurin and the opposition to DARPP-32 phosphorylation attenuates CREB phosphorylation. The net effect of the NMDA receptor pathway is a decrease of high forskolin-stimulated 133Ser-phospho-CREB levels. Therefore, inhibition of NMDA receptors increases CREB phosphorylation. The bold arrow points out the predominant action of the NMDA receptor pathway in the presence of high concentrations of forskolin.
Similar articles
-
Dopamine D2-like antagonists induce chromatin remodeling in striatal neurons through cyclic AMP-protein kinase A and NMDA receptor signaling.J Neurochem. 2004 Sep;90(5):1117-31. doi: 10.1111/j.1471-4159.2004.02569.x. J Neurochem. 2004. PMID: 15312167 Free PMC article.
-
The molecular basis of dopamine and glutamate interactions in the striatum.Adv Pharmacol. 1998;42:729-33. doi: 10.1016/s1054-3589(08)60851-0. Adv Pharmacol. 1998. PMID: 9328002 No abstract available.
-
Region-dependent dynamics of cAMP response element-binding protein phosphorylation in the basal ganglia.Proc Natl Acad Sci U S A. 1998 Apr 14;95(8):4708-13. doi: 10.1073/pnas.95.8.4708. Proc Natl Acad Sci U S A. 1998. PMID: 9539803 Free PMC article.
-
Food restriction increases NMDA receptor-mediated calcium-calmodulin kinase II and NMDA receptor/extracellular signal-regulated kinase 1/2-mediated cyclic amp response element-binding protein phosphorylation in nucleus accumbens upon D-1 dopamine receptor stimulation in rats.Neuroscience. 2005;132(4):1035-43. doi: 10.1016/j.neuroscience.2005.02.006. Neuroscience. 2005. PMID: 15857708
-
L-type Ca2+ channel blockers promote Ca2+ accumulation when dopamine receptors are activated in striatal neurons.Brain Res Mol Brain Res. 2004 Nov 24;131(1-2):65-72. doi: 10.1016/j.molbrainres.2004.08.007. Brain Res Mol Brain Res. 2004. PMID: 15530653 Free PMC article.
Cited by
-
Drug-induced activation of dopamine D(1) receptor signaling and inhibition of class I/II histone deacetylase induce chromatin remodeling in reward circuitry and modulate cocaine-related behaviors.Neuropsychopharmacology. 2008 Nov;33(12):2981-92. doi: 10.1038/npp.2008.15. Epub 2008 Feb 20. Neuropsychopharmacology. 2008. PMID: 18288092 Free PMC article.
-
L-Type Ca(2+) channels are essential for glutamate-mediated CREB phosphorylation and c-fos gene expression in striatal neurons.J Neurosci. 1999 Aug 1;19(15):6348-59. doi: 10.1523/JNEUROSCI.19-15-06348.1999. J Neurosci. 1999. PMID: 10414964 Free PMC article.
-
Expression and function of dopamine receptors in the developing medial frontal cortex and striatum of the rat.Neuroscience. 2011 Dec 29;199:501-14. doi: 10.1016/j.neuroscience.2011.10.004. Epub 2011 Oct 8. Neuroscience. 2011. PMID: 22015925 Free PMC article.
-
Dopamine D2-like antagonists induce chromatin remodeling in striatal neurons through cyclic AMP-protein kinase A and NMDA receptor signaling.J Neurochem. 2004 Sep;90(5):1117-31. doi: 10.1111/j.1471-4159.2004.02569.x. J Neurochem. 2004. PMID: 15312167 Free PMC article.
-
Psychostimulants, Protein phosphorylation and Gene expression: a growing role of L-type calcium channels.Cellscience. 2005 Jul 29;2(1):127-144. doi: 10.1901/jaba.2005.2-127. Cellscience. 2005. PMID: 16724157 Free PMC article. No abstract available.
References
-
- Matthies H. In search of cellular mechanisms of memory. Prog Neurobiol. 1989;32:277–349. - PubMed
-
- Deadwyler SA, Dunwiddie T, Lynch G. A critical level of protein synthesis is required for long-term potentiation. Synapse. 1987;1:90–95. - PubMed
-
- Schulman H. Protein phosphorylation in neuronal plasticity and gene expression. Curr Opin Neurobiol. 1995;5:375–381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
