

Review
doi: 10.1091/mbc.9.8.1951.
The ADF homology (ADF-H) domain: a highly exploited actin-binding module
Affiliations
- PMID: 9693358
- PMCID: PMC25446
- DOI: 10.1091/mbc.9.8.1951
Item in Clipboard
Review
The ADF homology (ADF-H) domain: a highly exploited actin-binding module
Mol Biol Cell.
1998 Aug.
Free PMC article
No abstract available
Figures
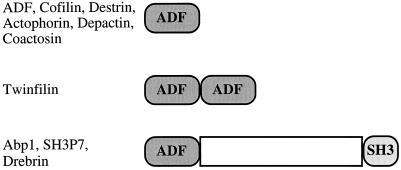
Domain structures of the three classes of ADF-H
domain proteins. The ADF/cofilin proteins are composed of a single
ADF-H domain. Twinfilins are composed of two ADF-H domains arranged in
tandem. Members of the drebrin/Abp1 class have an ADF-H domain at the
N-terminus of the protein, followed by a variable region and a
C-terminal SH3 domain.
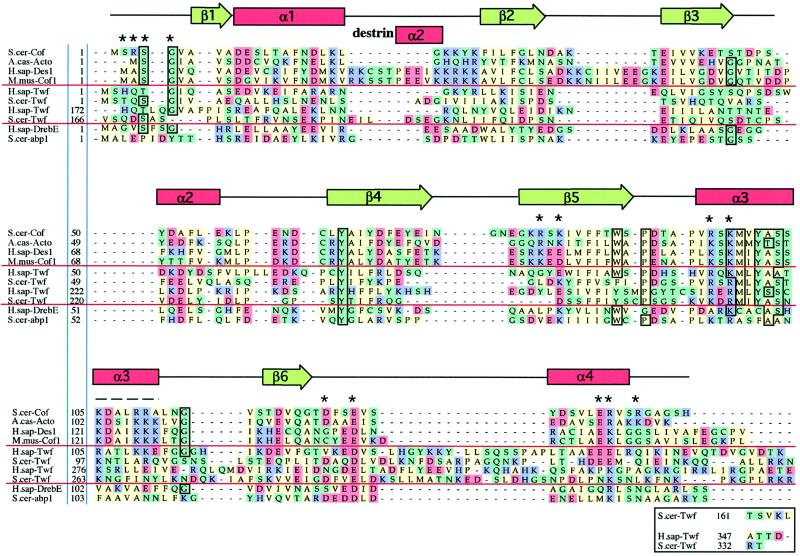
Sequence alignments of representative examples of
ADF-H domains. This alignment was produced essentially according to the
methods described for myosin motor domains by Cope et
al., (1996). Briefly, the collected sequences were subjected to
an initial alignment using the Clustal-W program (Thompson et
al., 1994). This initial alignment was refined, thus defining a
core ADF-H domain. Acidic (D and E), basic (K, R, and H), uncharged
nonpolar (A, I, M, V, L, F, W, and P), and other residues (Y, T, S, G,
N, Q, and C) have been colored in red, purple, green, and yellow,
respectively. The residues that are >75% conserved throughout the
entire ADF-H domain family are boxed. Dashes indicate positions
occupied by residues from other ADF/cofilin proteins within the full
alignment. The residues shown to be essential for interactions of yeast
cofilin with actin (Lappalainen et al., 1997) are
indicated by asterisks above the sequences, and the region that has
been shown to be important for actin interactions by peptide inhibition
studies (Yonezawa et al., 1989) is shown by a dashed
line above the sequences. The positions of secondary structure elements
based on the yeast cofilin crystal structure (Fedorov et
al., 1997) and the nuclear magnetic resonance structure of
human destrin (Hatanaka et al., 1996) are shown above
the sequences. Protein names, database, and accession numbers for the
sequences, respectively, are listed in the legend to Figure 3.
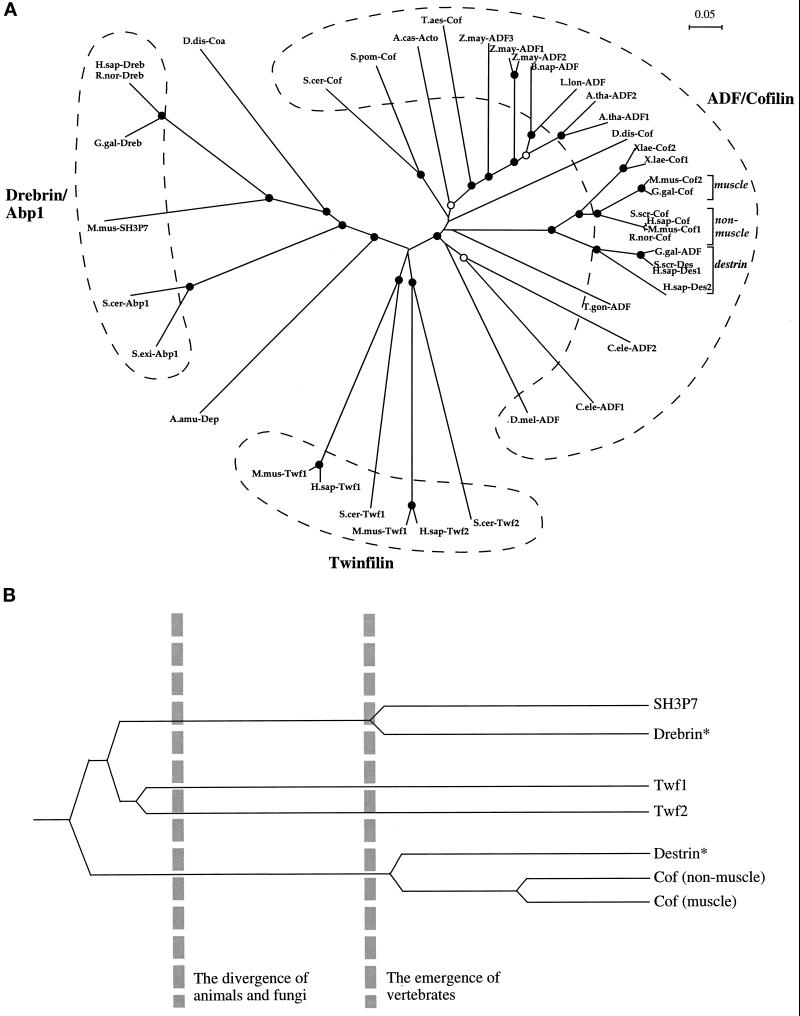
(A) An unrooted
phylogenetic tree of ADF-H domains. This tree was produced by
subjecting the alignment depicted in Figure 2 to analysis by the
Clustal-W software package (Thompson et al., 1994). An
allowance was made for multiple substitutions (Kimura, 1983).
Information from intervals in the alignment for which gaps are found in
some sequences was included. (Note: the tree architecture is almost
identical if gaps are omitted.) The tree was tested (1000 trials) for
branching order confidence by bootstrapping (Felsenstein, 1985). Filled
circles indicate branch points supported beyond a confidence level of
85%. Empty circles indicate branch points supported beyond the 50%
but below the 85% confidence level. Dashed lines indicate the three
classes described in this essay. A bar showing 5% divergence is
included. Further information on this procedure (as applied to myosin
motor domains) can be found on the Worldwide Web at
http://www.mrc-lmb.cam.ac.uk/ . (B) A simplified, rooted tree depicting
the evolution of ADF-H domain proteins in mice. All three families of
ADF-H domain proteins were present in the common ancestor of yeast and
animals. Distinct members of the ADF/cofilin family in mouse arose
after the emergence of vertebrates. The asterisks denote our
predictions that destrin- and drebrin-like proteins will be found in
mouse, based on the phylogenetic tree shown in A. Protein names,
database, and accession numbers for the sequences, respectively, are
listed below. Where no database is stated, the accession number refers
to GenBank. S. cerevisiae cofilin: Swiss-Prot, Q03048;
S. pombe cofilin: DDBJ, D89939; D.
discoideum cofilin: Swiss-Prot, P54706; A.
castellanii actophorin: Swiss-Prot, P37167; A.
thaliana ADF1: U48938; A. thaliana ADF2: U48939;
L. longifolium ADF: PIR, S30935; Brassica
napus ADF: PIR, S30934; Z. mays ADF1:
Swiss-Prot, P46251; Z. mays ADF2: X97725; Z.
mays ADF3: X97726; Triticum aestivum cofilin:
U58278; Drosophila melanogaster ADF: PIR, A57569;
Caenorhabditis elegans ADF1: Swiss-Prot, Q07750;
C. elegans ADF2 (Swiss-Prot: Q07749), T.
gondii ADF: U62146; H. sapiens destrin
2:(U47924; H. sapiens destrin 1: PIR, A54184; S.
scrofa destrin: DDBJ, D90053; Gallus gallus ADF:
J02912; R. norvegicus cofilin: Swiss-Prot, P45592;
Mus musculus cofilin (nonmuscle isoform): Swiss-Prot,
P18760; H. sapiens cofilin: EMBL, X95404; S.
scrofa cofilin: M20866; G. gallus cofilin:
M55659; M. musculus cofilin (muscle isoform):
Swiss-Prot, P45591; X. laevis cofilin 1: U26270;
X. laevis cofilin 2: Swiss-Prot, P45593; M.
musculus twinfilin (repeat-1): U82324; H.
sapiens twinfilin (repeat-1): PIR, A55922; S.
cerevisiae twinfilin (repeat-1): SGD, YGR080W; M.
musculus twinfilin (repeat-2): U82324; H.
sapiens twinfilin (repeat-2): PIR, 55922; S.
cerevisiae twinfilin (repeat-2): SGD, YGR080W; H.
sapiens drebrin E: Swiss-Prot, Q16643; R.
norvegicus drebrin A: Swiss-Prot, Q07266; G.
gallus drebrin A, E1 and E2: Swiss-Prot, P18302; M.
musculus SH3P7: GenBank, U58884; D.
discoideum coactosin: Swiss-Prot, P34121; S.
cerevisiae Abp1: EMBL, X51780/Swiss-Prot, P15891; S.
exiguus Abp1: Swiss-Prot, P38479; A. amurensis
depactin: Swiss-Prot, P20690.
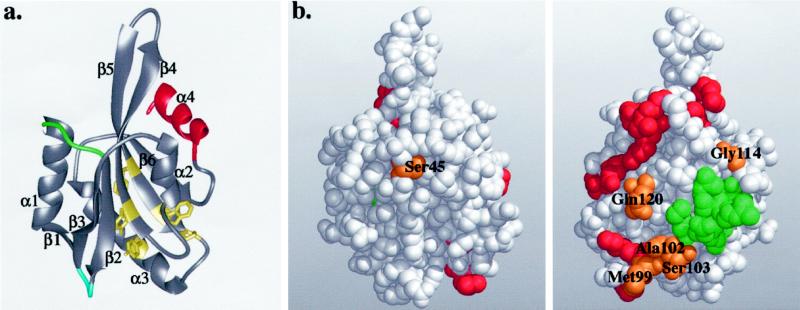
(a) Ribbon diagram of the yeast cofilin structure.
Cofilin has a central mixed β-sheet, which is sandwiched between two
pairs of α-helices. The positions of the insertions in mammalian
cofilins are in green and blue, and the diverged region in twinfilins
is in red. The highly conserved residues that appear to be important
for protein stability and correct folding (Tyr64,
Phe85, Trp88, Pro90, and
Tyr101) are shown in yellow. (b) Space-filling model of
yeast cofilin shown in two different orientations (rotated 180°
around the y-axis). The residues that are essential for actin
interactions in yeast cofilin are highlighted in red. The residues
implicated in actin binding by peptide inhibition studies are in green.
The highly conserved surface residues (Ser45,
Met99, Ala102, Ser103,
Gly114, and Gln120) are in orange. All of these
residues, except Ser45, are located close to site of
cofilin implicated genetically in actin binding, suggesting they also
form part of the actin-binding surface. These Figures were produced
using Midas Software (University of California San Francisco) running
on a Silicon Graphics (Mountain View, CA) Indigo II workstation.
References
-
- Agnew BJ, Minamide LS, Bamburg JR. Reactivation of phosphorylated actin depolymerizing factor and identification of the regulatory site. J Biol Chem. 1995;270:17582–17587. - PubMed
-
- Bernstein BW, Bamburg JR. Tropomyosin binding to F-actin protects the F-actin from disassembly by brain actin-depolymerizing-factor (ADF) Cell Motil. 1982;2:1–8. - PubMed
-
- Cope MJTV, Whisstock J, Rayment I, Kendrick-Jones J. Conservation within the myosin motor domain—implications for structure and function. Structure. 1996;15:969–987. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
