

Cytokinesis and midzone microtubule organization in Caenorhabditis elegans require the kinesin-like protein ZEN-4
- PMID: 9693365
- PMCID: PMC25457
- DOI: 10.1091/mbc.9.8.2037
Cytokinesis and midzone microtubule organization in Caenorhabditis elegans require the kinesin-like protein ZEN-4
Abstract
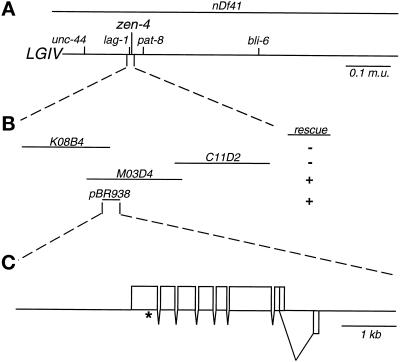
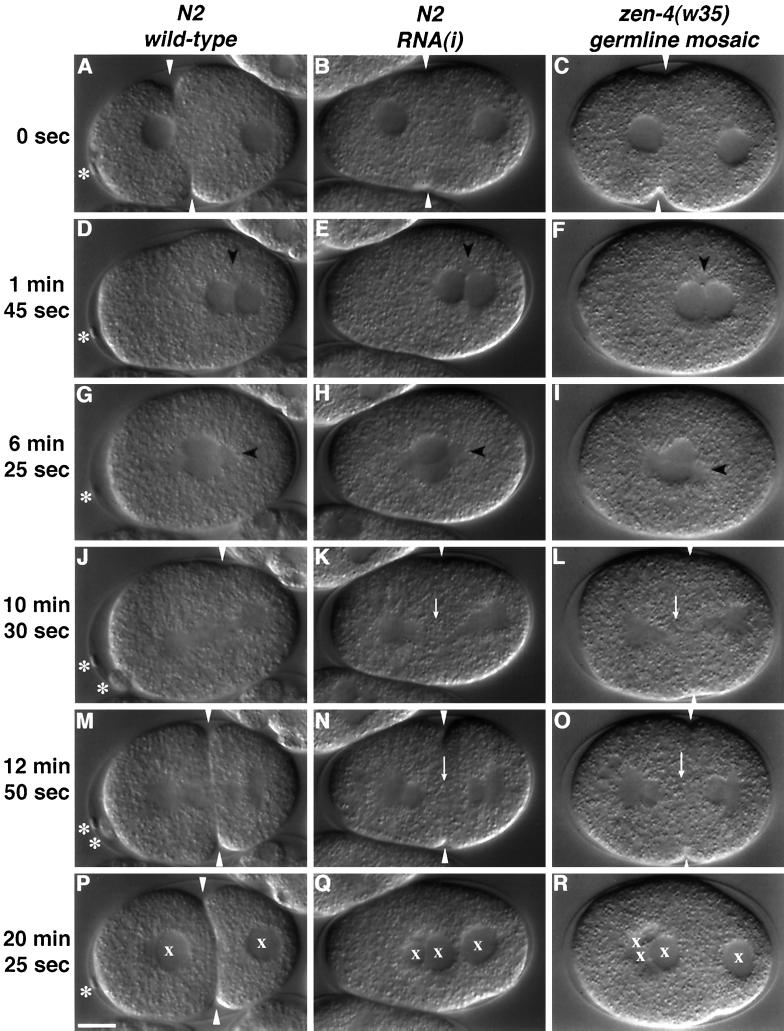
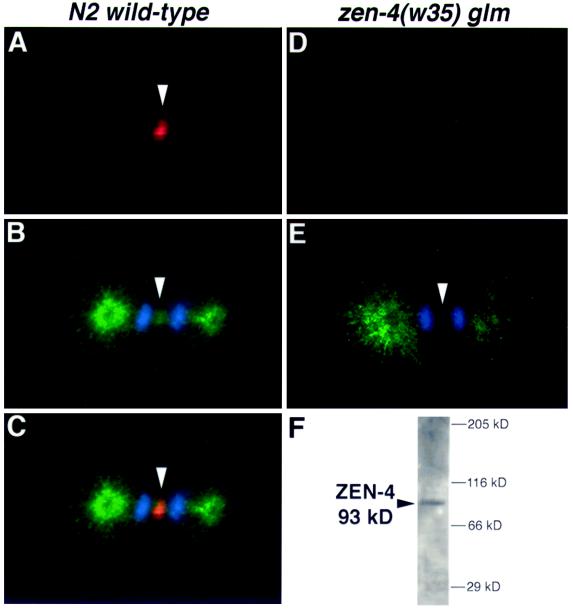
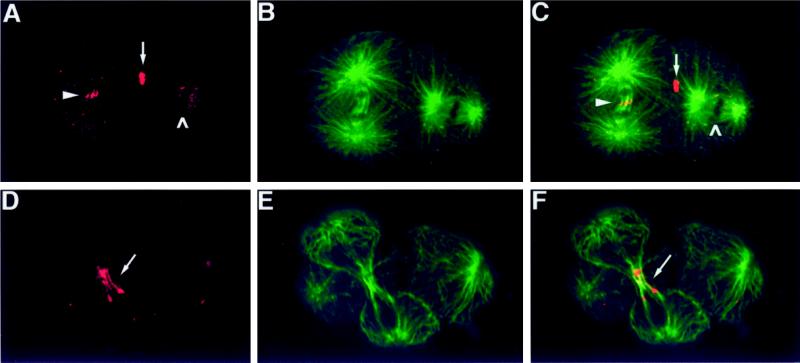
Members of the MKLP1 subfamily of kinesin motor proteins localize to the equatorial region of the spindle midzone and are capable of bundling antiparallel microtubules in vitro. Despite these intriguing characteristics, it is unclear what role these kinesins play in dividing cells, particularly within the context of a developing embryo. Here, we report the identification of a null allele of zen-4, an MKLP1 homologue in the nematode Caenorhabditis elegans, and demonstrate that ZEN-4 is essential for cytokinesis. Embryos deprived of ZEN-4 form multinucleate single-celled embryos as they continue to cycle through mitosis but fail to complete cell division. Initiation of the cytokinetic furrow occurs at the normal time and place, but furrow propagation halts prematurely. Time-lapse recordings and microtubule staining reveal that the cytokinesis defect is preceded by the dissociation of the midzone microtubules. We show that ZEN-4 protein localizes to the spindle midzone during anaphase and persists at the midbody region throughout cytokinesis. We propose that ZEN-4 directly cross-links the midzone microtubules and suggest that these microtubules are required for the completion of cytokinesis.
Figures


Comment in
-
An MBoC favorite: Cytokinesis and midzone microtubule organization in Caenorhabditis elegans require the kinesin-like protein ZEN-4.Mol Biol Cell. 2012 Aug;23(16):3025. doi: 10.1091/mbc.E12-02-0149. Mol Biol Cell. 2012. PMID: 22891030 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
