

doi: 10.1101/gad.12.15.2434.
E1A signaling to p53 involves the p19(ARF) tumor suppressor
Affiliations
- PMID: 9694807
- PMCID: PMC317046
- DOI: 10.1101/gad.12.15.2434
Item in Clipboard
E1A signaling to p53 involves the p19(ARF) tumor suppressor
Genes Dev.
.
Abstract
The adenovirus E1A oncogene activates p53 through a signaling pathway involving the retinoblastoma protein and the tumor suppressor p19(ARF). The ability of E1A to induce p53 and its transcriptional targets is severely compromised in ARF-null cells, which remain resistant to apoptosis following serum depletion or adriamycin treatment. Reintroduction of p19(ARF) restores p53 accumulation and resensitizes ARF-null cells to apoptotic signals. Therefore, p19(ARF) functions as part of a p53-dependent failsafe mechanism to counter uncontrolled proliferation. Synergistic effects between the p19(ARF) and DNA damage pathways in inducing p53 may contribute to E1A's ability to enhance radio- and chemosensitivity.
Figures
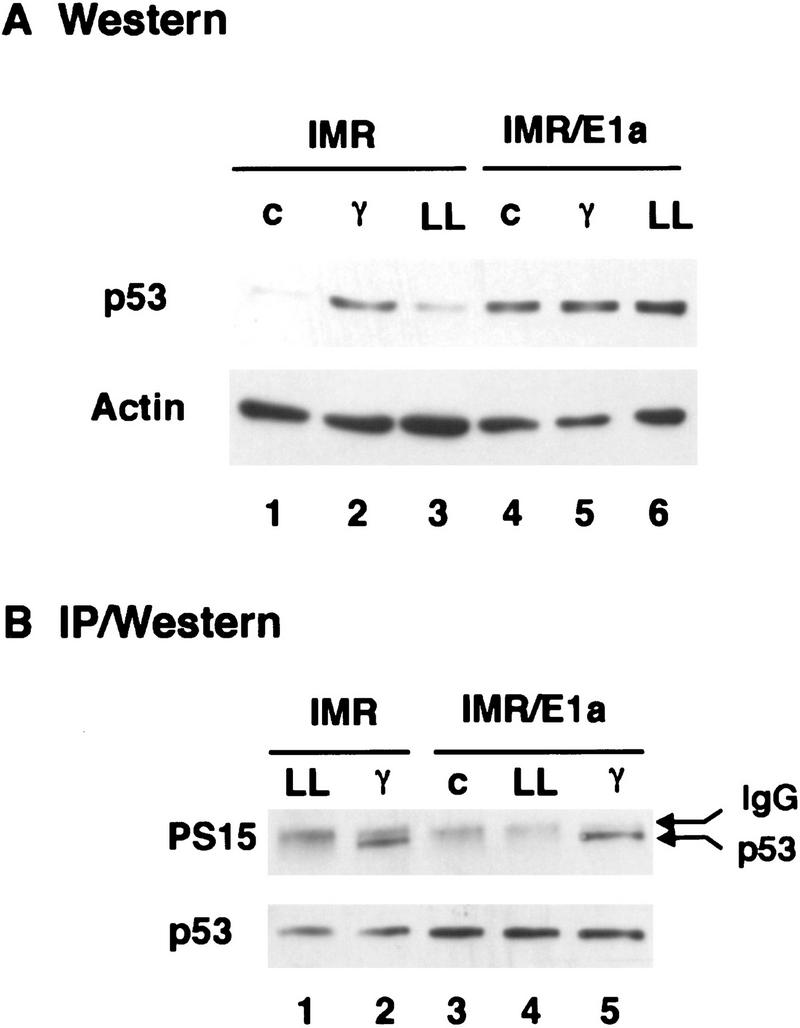
E1A induces p53 in the absence of phosphorylation on serine-15. IMR90 fibroblasts were infected with control (IMR) or E1A-expressing (IMR/E1A) retroviruses. Extracts were prepared from untreated cells (c), or from cells treated 3 hr earlier with 7 Gy γ radiation (γ) or 2 hr earlier with 50 μm LLnL (LL). (A) p53 levels were determined by Western blot analysis using pAb 1801 and DO1. Equal loading of the gel was confirmed by stripping the blot and reprobing with anti-β-actin antiserum. (B) p53 was immunoprecipitated from extracts corresponding to 100 μg (IMR) or 35 μg (IMR/E1A) total protein using pAb 1801, and Western blots were probed with antibodies specific for p53 phosphoserine-15 (αp53-P–Ser-15).
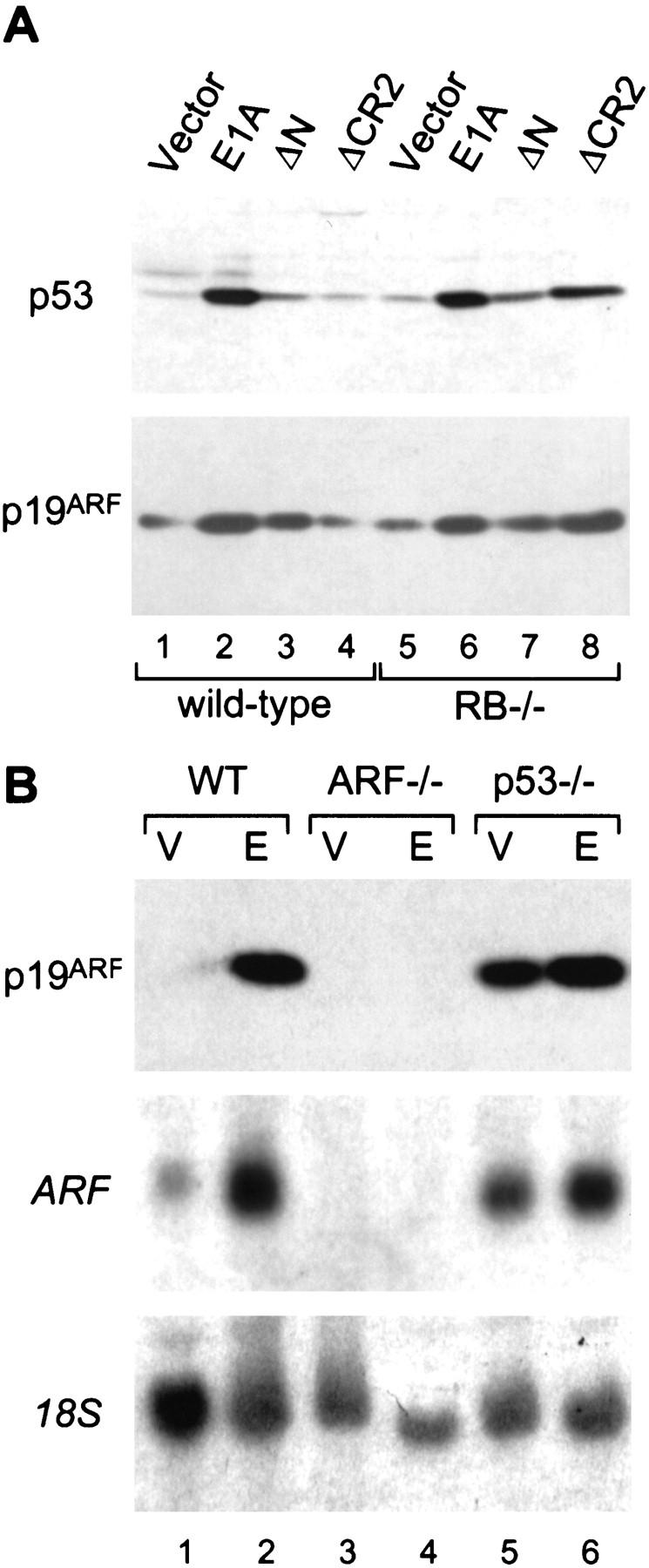
E1A induces p19ARF and p53 through a similar mechanism. (A) Early passage (about three to four) wild-type and Rb−/− MEFs from littermates embryos were infected with retroviruses expressing full-length E1A or E1A mutants unable to bind p300/CBP (ΔN) or the Rb-related proteins (ΔCR2). An empty retroviral vector was used as a control (vector). Immunoblotting was performed using polyclonal antibodies against p19ARF or p53. Using this procedure, each E1A mutant is efficiently expressed at comparable levels (Samuelson et al. 1997). (B) Wild-type (WT), ARF-null (ARF−/−), and p53-null (p53−/−) MEFs were infected with a control vector (V) or a retrovirus expressing full-length E1A (E). Lysates were derived from whole populations passaged minimally in culture (<1 week) and analyzed for ARF protein (top) or mRNA (middle) expression by Western or Northern blotting, respectively. Northern blots were rehybridized using a probe to the 18S rRNA to confirm equal loading (bottom).
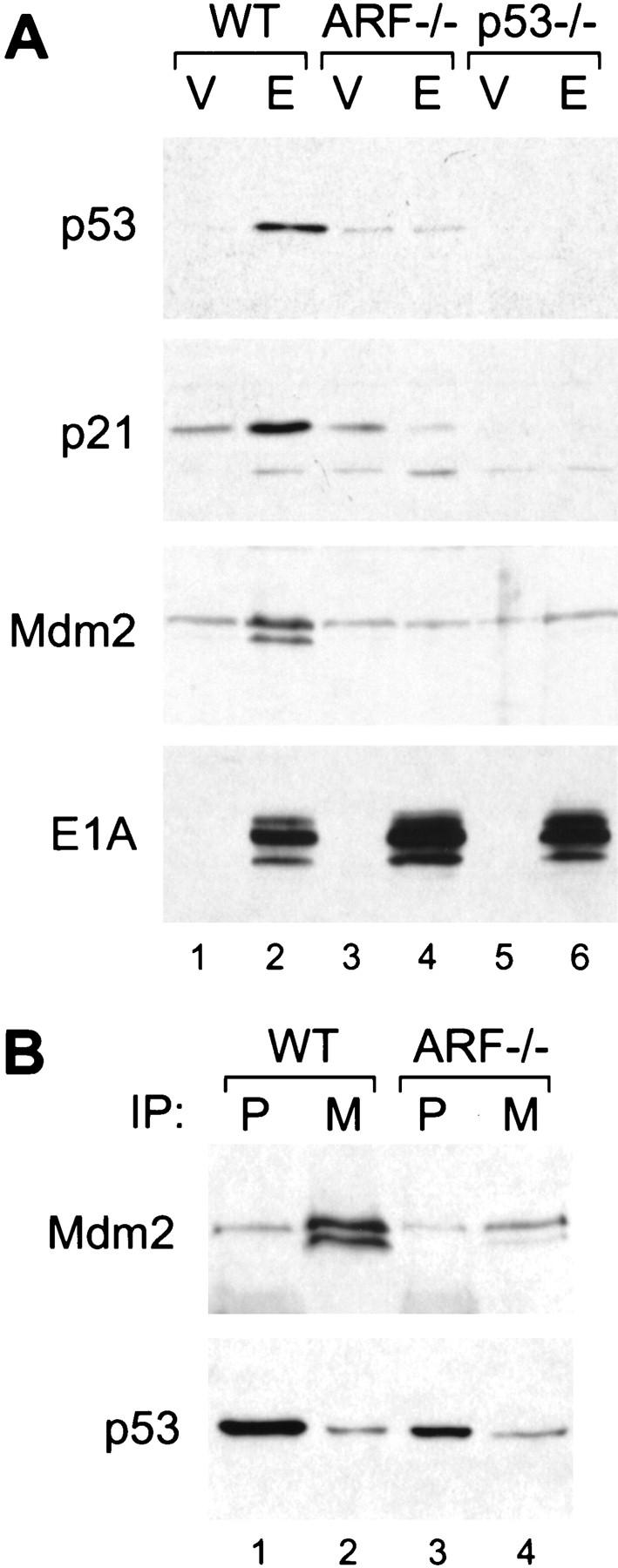
p19ARF mediates p53 induction by E1A and interferes with the p53/Mdm2 interaction. Wild-type (WT), ARF-null (ARF−/−) and p53-null (p53−/−) cell populations harboring a control vector (V) or expressing E1A (E) were prepared by retroviral gene transfer. Protein expression was analyzed in whole cell populations passaged minimally in culture (<1 week). (A) p53 protein levels along with the levels of its transcriptional targets p21 and Mdm2 were determined by immunoblotting. (B) Mdm2/p53 complexes were examined in wild-type and ARF-null populations expressing E1A by immunoprecipitation with monoclonal antibodies directed against p53 (P) or Mdm2 (M), followed by immunoblotting with a polyclonal rabbit antibody against p53. The blots were then reprobed using the same monoclonal antibody against Mdm2. Note that the p53 blot was overexposed to allow visualization of the amount associated with Mdm2.
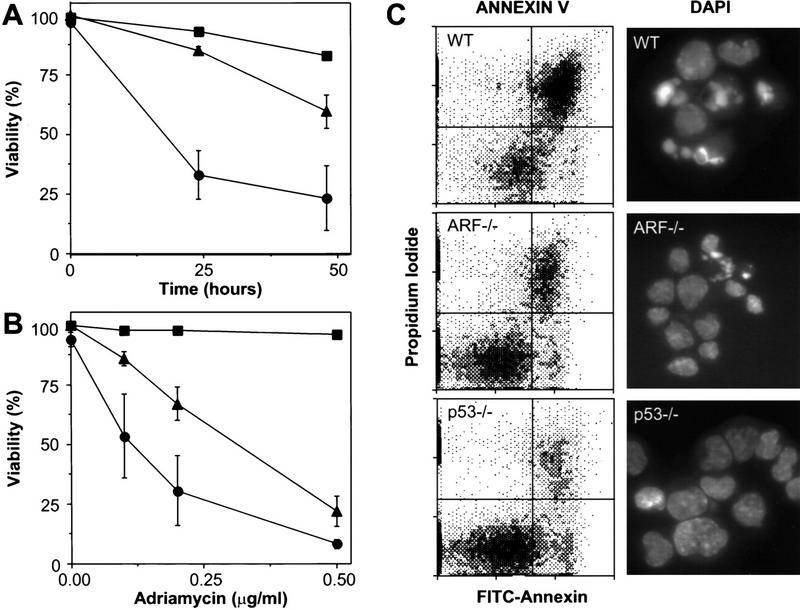
E1A-expressing cells lacking ARF are defective in apoptosis. Wild-type (•), ARF-null (▴), and p53-null (▪) early passage MEFs were infected with control retroviruses (not shown) or retroviruses expressing E1A. Within a week of gene transfer, the resulting cell populations were examined for cell death at various times following serum depletion (A) or 24 hr after treatment with the indicated doses of adriamycin (B). Cell viability was assessed by trypan blue exclusion. Each point represents the mean±s.d. from at least three separate experiments. Fibroblasts of all genotypes infected with a control vector retained viability (>90%) following serum depletion or adriamycin treatment (data not shown). (C) Wild-type (WT), ARF-null (ARF−/−) and p53-null (p53−/−) MEFs expressing E1A were examined for apoptosis 18 hr after transfer to 0.1% serum conditions. Annexin V binds phosphotidylserine. Apoptotic changes in membrane biochemistry lead to increased concentration of phosphotidylserine on the outer plasma membrane, where it becomes accessible to annexin V (Andree et al. 1990). Propidium iodide fluorescently stains late apoptotic cells that have lost membrane integrity. Shown are representative dot plots from two-color flow cytometry: (Bottom left quadrant) Viable; (bottom right quadrant) early apoptotic; (top right quadrant) late apoptotic. DAPI staining allows visualization of the chromatin condensation characteristic of apoptotic cells. Note that there was little apoptosis in E1A-expressing populations in 10% serum nor in vector-only control populations in 0.1% serum (data not shown).
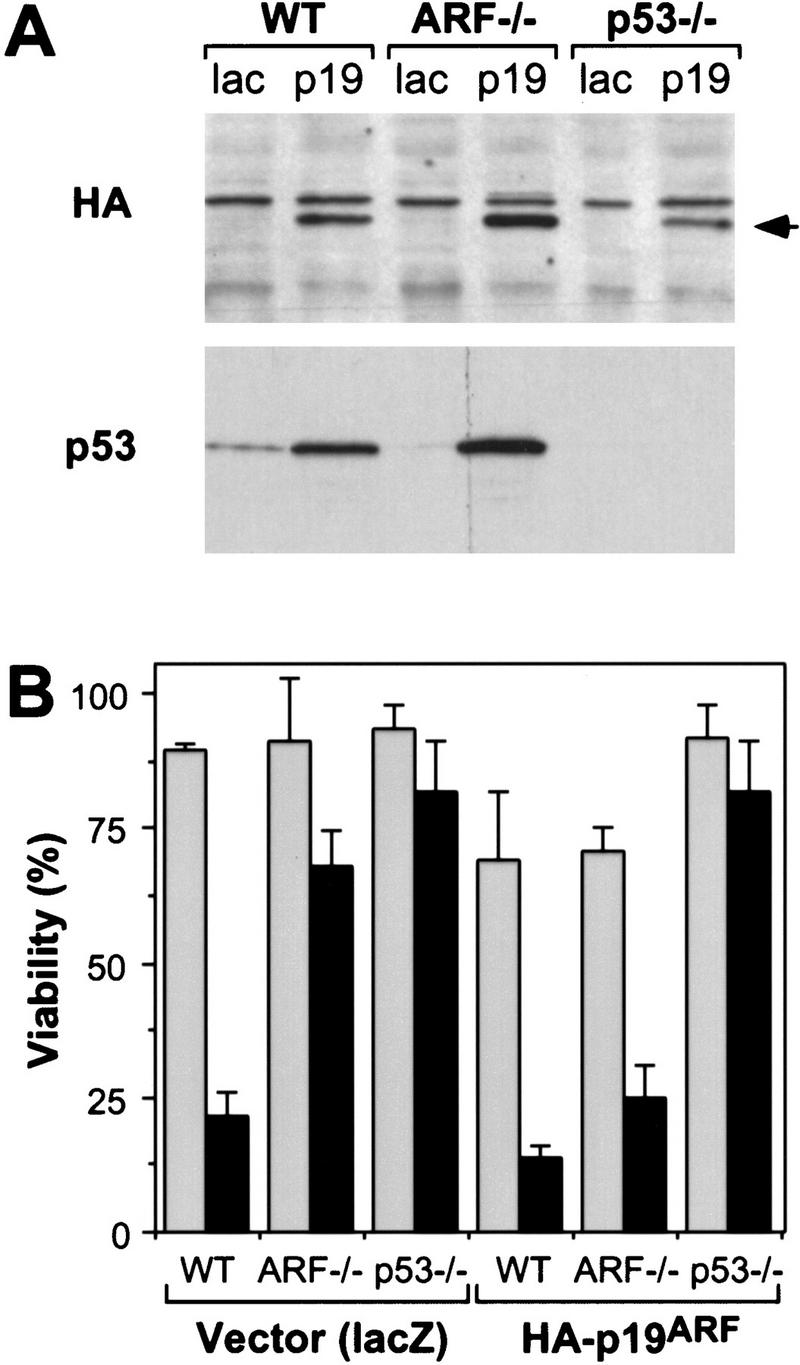
Reintroduction of p19ARF restores apoptosis. Control and E1A-expressing populations derived from wild-type (WT), ARF-null (ARF−/−) and p53-null (p53−/−) populations were infected with retroviruses expressing lacZ or an HA-tagged ARF cDNA (Quelle et al. 1995). Thirty-six hours later, the resulting cell populations were analyzed for p53 and exogenous p19ARF protein expression or treated with apoptotic stimuli. (A) Immunoblotting of infected populations using a monoclonal antibody recognizing the HA epitope fused to p19ARF or a polyclonal antibody directed against p53. The arrow denotes the migration of HA-tagged p19ARF. (B) The indicated cell populations were placed in 10% (shaded bars) or 0.1% (solid bars) serum for 24 hr and cell viability was measured by trypan blue exclusion. The values represent the mean and s.d. of at least three separate infections.
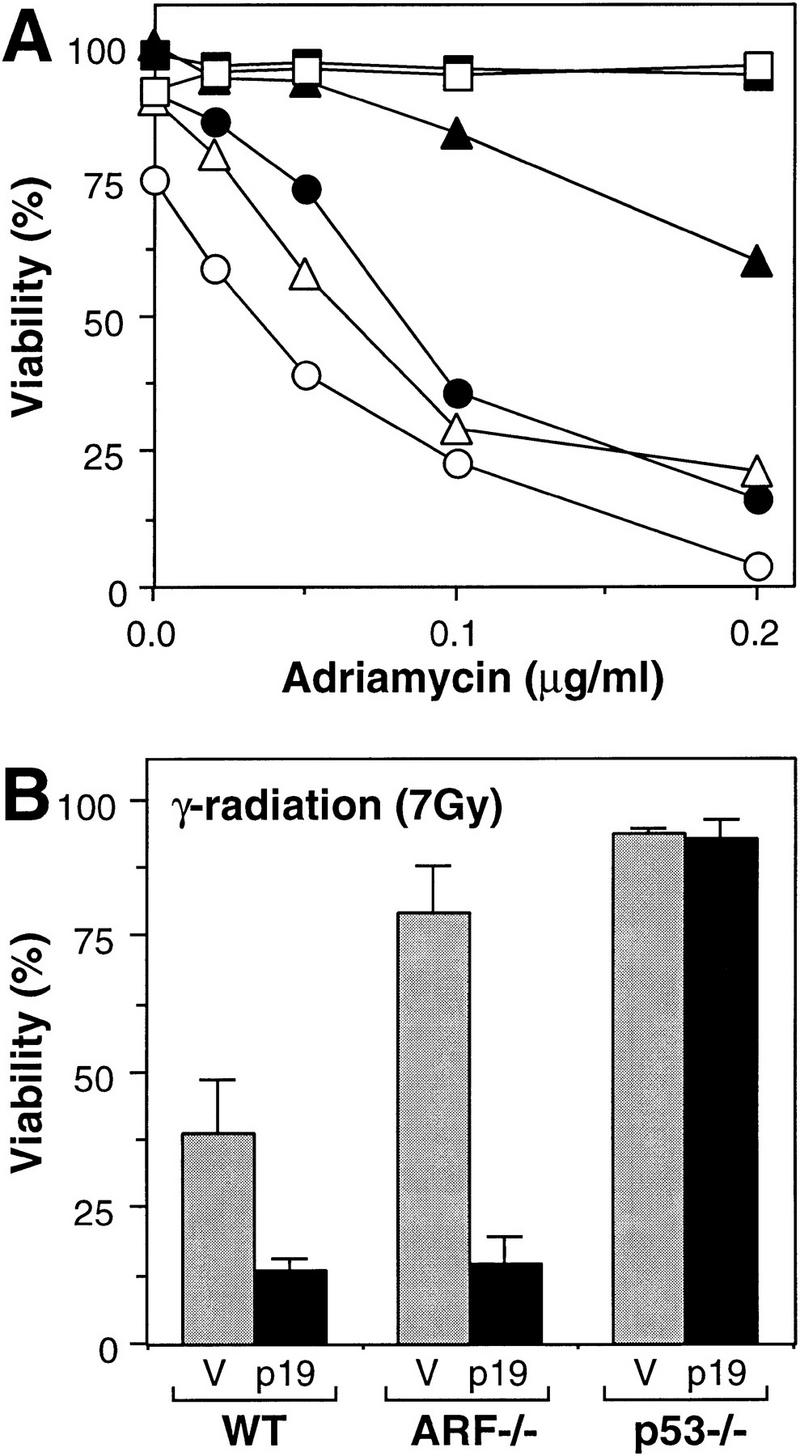
Synergy between p19ARF-dependent and -independent pathways targeting p53. (A) lacZ (solid symbols)- and HA–ARF (open symbols)-expressing cell populations were treated with the indicated doses of adriamycin, and cell viability was determined 24 hr later by trypan blue exclusion. The cell populations were as follows: Wild-type MEFs lacking E1A (squares); wild-type MEFs expressing E1A (circles); ARF−/− MEFs expressing E1A (triangles). Note that ARF−/− and p53−/− MEFs lacking E1A, as well as p53-deficient MEFs expressing E1A, remained viable in adriamycin whether or not they expressed HA–p19ARF (data not shown). (B) lacZ (V, shaded bars) and HA–p19ARF (p19, solid bars) expressing cell populations were treated with 7 Gy ionizing radiation and cell viability was determined 24 hr later by trypan blue exclusion. The values represent the mean and s.d. of at least three separate populations. MEFs not expressing E1A were resistant to apoptosis under these conditions (data not shown; see also Lowe et al. 1993).
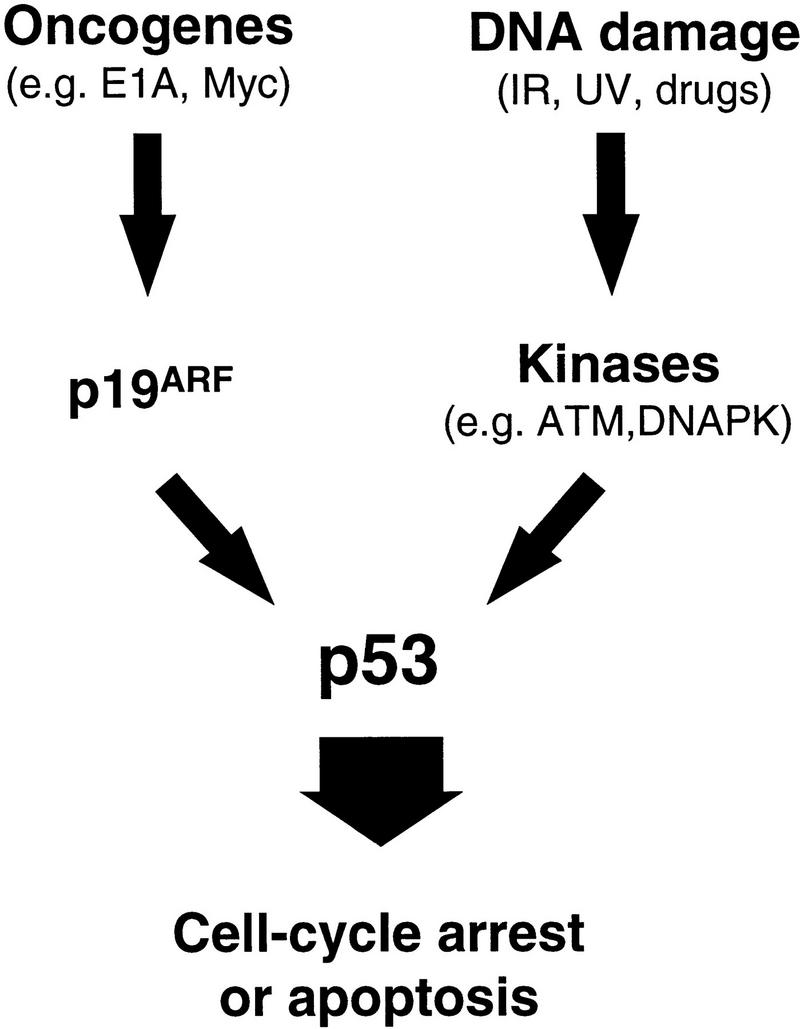
Oncogenes and DNA damage activate p53 through distinct mechanisms. p19ARF acts as an intermediary in p53 activation by mitogenic oncogenes such as E1A and myc. In contrast, activation of p53 following DNA damage involves de novo phosphorylation of p53 on serine-15 (and other residues) by kinases such as the DNA-dependent protein kinase (DNA–PK) or the product of the ataxia-telangiectasia gene (ATM) (Shieh et al. 1997; Siliciano et al. 1997). Activation of p53 by oncogenes does not involve phosphorylation on serine-15, and both serine-15 phosphorylation (not shown) and p53 activation (Kamijo et al. 1997) following DNA damage are unimpaired in the absence of ARF. Therefore, the two upstream signaling pathways to p53 are fundamentally distinct.
References
-
- Andree HA, Reutelingsperger CP, Hauptmann R, Hemker HC, Hermens WT, Willems GM. Binding of vascular anticoagulant alpha (VAC alpha) to planar phospholipid bilayers. J Biol Chem. 1990;265:4923–4928. - PubMed
-
- Barak Y, Gottlieb E, Juvengershon T, Oren M. Regulation of mdm2 expression by p53: Alternative promoters produce transcripts with nonidentical translation potential. Genes & Dev. 1994;8:1739–1749. - PubMed
-
- Debbas M, White E. Wild-type p53 mediates apoptosis by E1A, which is inhibited by E1B. Genes & Dev. 1993;7:546–554. - PubMed
-
- DeCaprio JA, Ludlow JW, Lynch D, Furukawa Y, Griffin J, Piwnica-Worms H, Huang CM, Livingstone DM. SV40 large T antigen forms a specific complex with the product of the retinoblastoma susceptibility gene. Cell. 1988;54:275–283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous