

Interactions between the structural domains of the RNA replication proteins of plant-infecting RNA viruses
- PMID: 9696810
- PMCID: PMC109938
- DOI: 10.1128/JVI.72.9.7160-7169.1998
Interactions between the structural domains of the RNA replication proteins of plant-infecting RNA viruses
Abstract
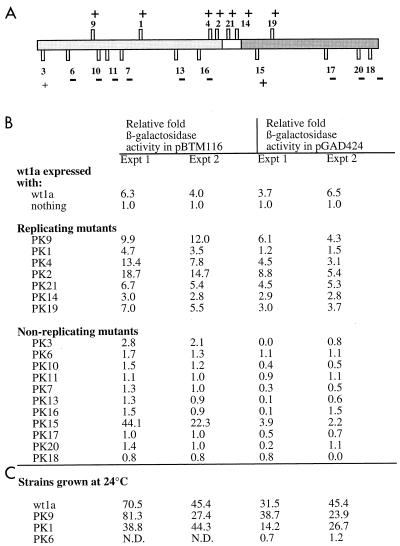
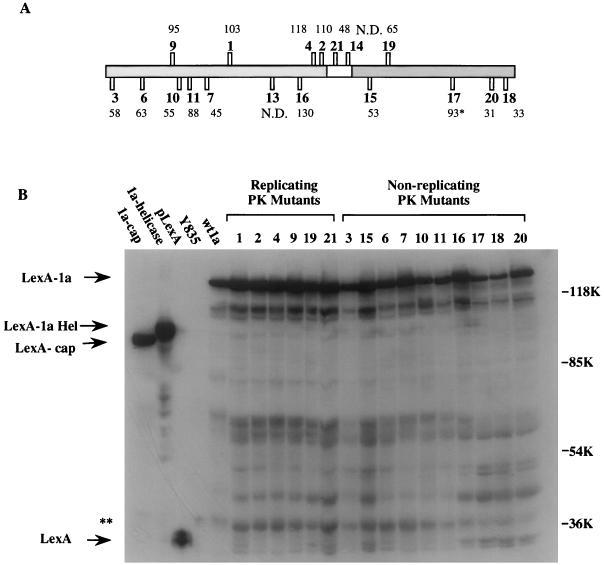
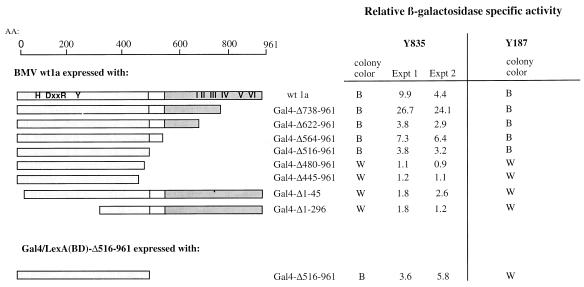
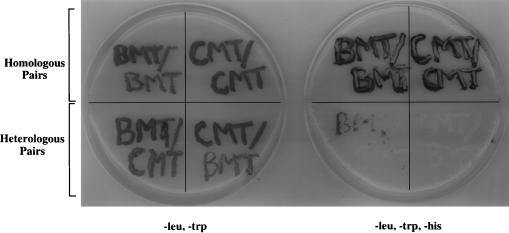
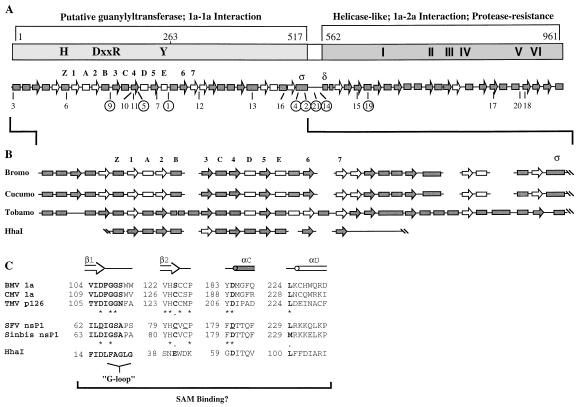
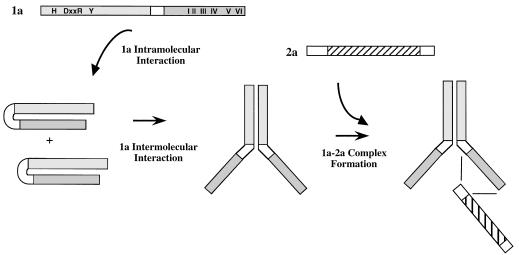
Brome mosaic virus (BMV), a positive-strand RNA virus, encodes two replication proteins: the 2a protein, which contains polymerase-like sequences, and the 1a protein, with N-terminal putative capping and C-terminal helicase-like sequences. These two proteins are part of a multisubunit complex which is necessary for viral RNA replication. We have previously shown that the yeast two-hybrid assay consistently duplicated results obtained from in vivo RNA replication assays and biochemical assays of protein-protein interaction, thus permitting the identification of additional interacting domains. We now map an interaction found to take place between two 1a proteins. Using previously characterized 1a mutants, a perfect correlation was found between the in vivo phenotypes of these mutants and their abilities to interact with wild-type 1a (wt1a) and each other. Western blot analysis revealed that the stabilities of many of the noninteracting mutant proteins were similar to that of wt1a. Deletion analysis of 1a revealed that the N-terminal 515 residues of the 1a protein are required and sufficient for 1a-1a interaction. This intermolecular interaction between the putative capping domain and itself was detected in another tripartite RNA virus, cucumber mosaic virus (CMV), suggesting that the 1a-1a interaction is a feature necessary for the replication of tripartite RNA viruses. The boundaries for various activities are placed in the context of the predicted secondary structures of several 1a-like proteins of members of the alphavirus-like superfamily. Additionally, we found a novel interaction between the putative capping and helicase-like portions of the BMV and CMV 1a proteins. Our cumulative data suggest a working model for the assembly of the BMV RNA replicase.
Figures
References
-
- Blumenthal T, Carmichael G. RNA replication: function and structure of Qβ-replicase. Annu Rev Biochem. 1979;48:525–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
