

Measles virus spread and pathogenesis in genetically modified mice
- PMID: 9696838
- PMCID: PMC109970
- DOI: 10.1128/JVI.72.9.7420-7427.1998
Measles virus spread and pathogenesis in genetically modified mice
Abstract
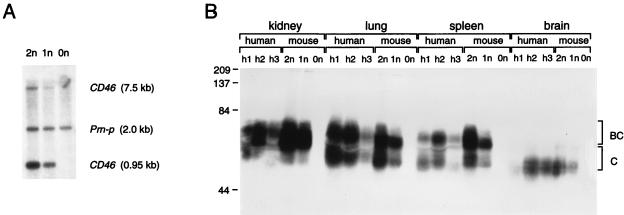
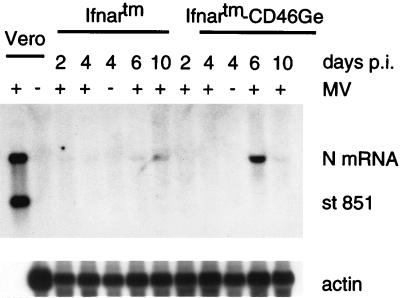
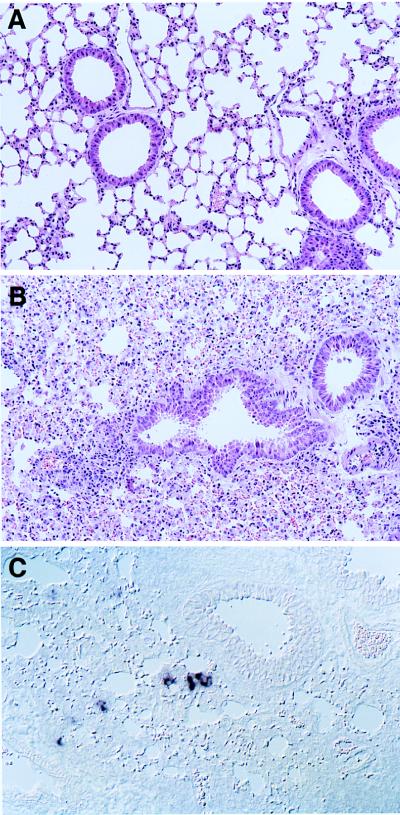
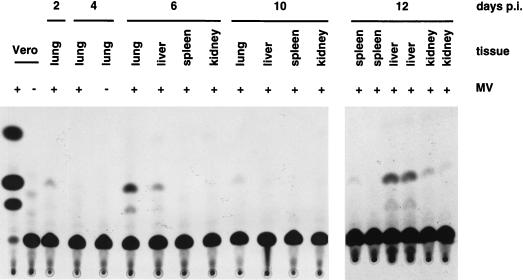
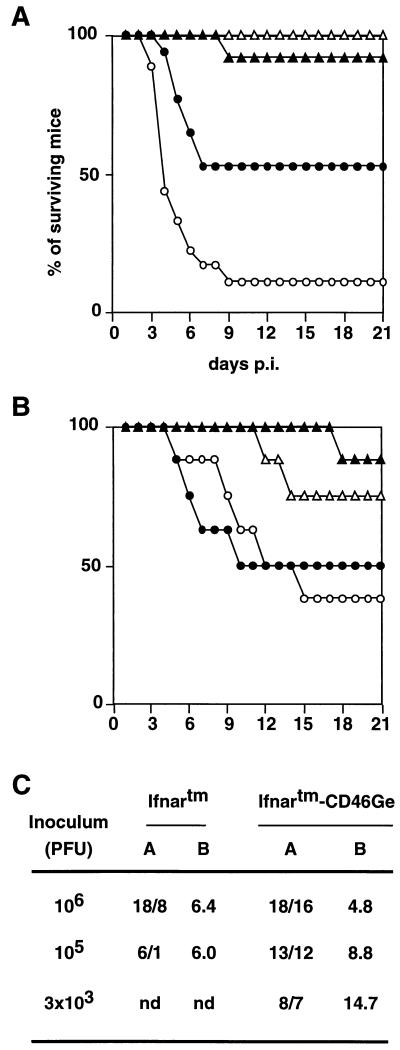
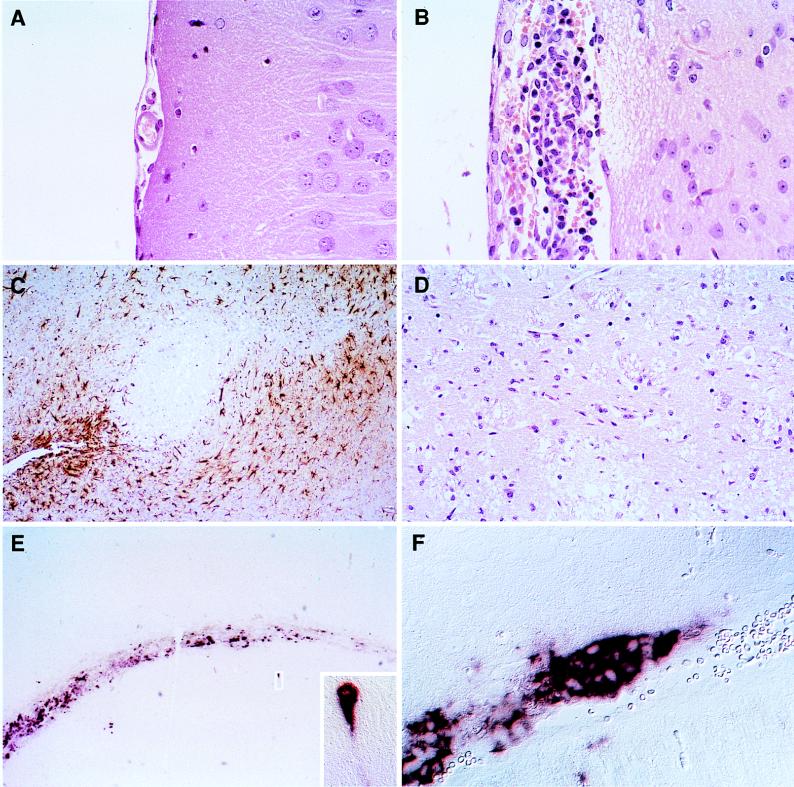
Attenuated Edmonston measles virus (MV-Edm) is not pathogenic in standard mice. We show here that MV-Edm inoculated via the natural respiratory route has a limited propagation in the lungs of mice with a targeted mutation inactivating the alpha/beta interferon receptor. A high dose of MV-Edm administered intracerebrally is lethal for about half of these mice. To study the consequences of the availability of a high-affinity receptor for MV propagation, we generated alpha/beta interferon-defective mice expressing human CD46 with human-like tissue specificity. Intranasal infection of these mice with MV-Edm resulted in enhanced spread to the lungs and more prominent inflammatory response. Virus replication was also detected in peripheral blood mononuclear cells, the spleen, and the liver. Moreover, intracerebral inoculation of adult animals with low MV-Edm doses caused encephalitis with almost inevitably lethal outcome. We conclude that in mice alpha/beta interferon controls MV infection and that a high-affinity receptor facilitates, but is not strictly required for, MV spread and pathogenesis.
Figures
References
-
- Buchholz C J, Gerlier D, Hu A, Cathomen T, Liszewski M K, Atkinson J P, Cattaneo R. Selective expression of a subset of measles virus receptor-competent CD46 isoforms in human brain. Virology. 1996;217:349–355. - PubMed
-
- Buchholz C J, Koller D, Devaux P, Mumenthaler C, Schneider-Schaulies J, Braun W, Gerlier D, Cattaneo R. Mapping of the primary binding site of measles virus to its receptor CD46. J Biol Chem. 1997;272:22072–22079. - PubMed
-
- Bueler H, Fischer M, Lang Y, Bluethmann H, Lipp H P, DeArmond S J, Prusiner S B, Aguet M, Weissmann C. Normal development and behaviour of mice lacking the neuronal cell-surface PrP protein. Nature. 1992;356:577–582. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
