

The role of influenza A virus hemagglutinin residues 226 and 228 in receptor specificity and host range restriction
- PMID: 9696865
- PMCID: PMC110023
- DOI: 10.1128/JVI.72.9.7626-7631.1998
The role of influenza A virus hemagglutinin residues 226 and 228 in receptor specificity and host range restriction
Abstract
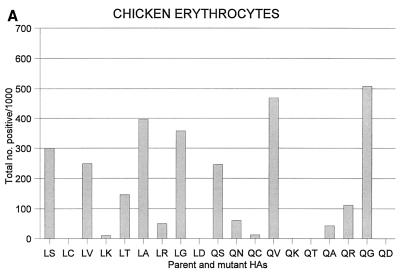
Influenza A viruses can be isolated from a variety of animals, but their range of hosts is restricted. For example, human influenza viruses do not replicate in duck intestine, the major replication site of avian viruses in ducks. Although amino acids at positions 226 and 228 of hemagglutinin (HA) of the H3 subtype are known to be important for this host range restriction, the contributions of specific amino acids at these positions to restriction were not known. Here, we address this issue by generating HAs with site-specific mutations of a human virus that contain different amino acid residues at these positions. We also let ducks select replication-competent viruses from a replication-incompetent virus containing a human virus HA by inoculating animals with 10(10.5) 50% egg infectious dose of the latter virus and identified a mutation in the HA. Our results showed that the Ser-to-Gly mutation at position 228, in addition to the Leu-to-Gln mutation at position 226 of the HA of the H3 subtype, is critical for human virus HA to support virus replication in duck intestine.
Figures
References
-
- Almond J W. A single gene determines the host range of influenza virus. Nature. 1977;270:617–618. - PubMed
-
- Beare A S, Webster R G. Replication of avian influenza viruses in humans. Arch Virol. 1991;119:37–42. - PubMed
-
- Connor R J, Kawaoka Y, Webster R G, Paulson J C. Receptor specificity in human, avian, and equine H2 and H3 influenza virus isolates. Virology. 1994;205:17–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
