

Immune complex-like moieties in immunoglobulin for intravenous use (i.v.Ig) bind complement and enhance phagocytosis of human erythrocytes
- PMID: 9697987
- PMCID: PMC1905024
- DOI: 10.1046/j.1365-2249.1998.00624.x
Immune complex-like moieties in immunoglobulin for intravenous use (i.v.Ig) bind complement and enhance phagocytosis of human erythrocytes
Abstract
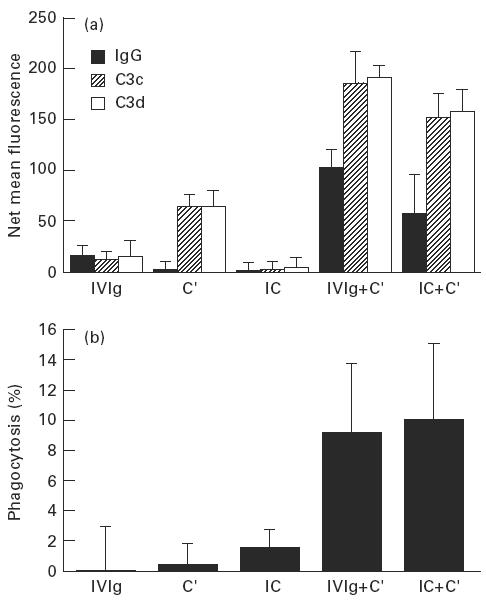
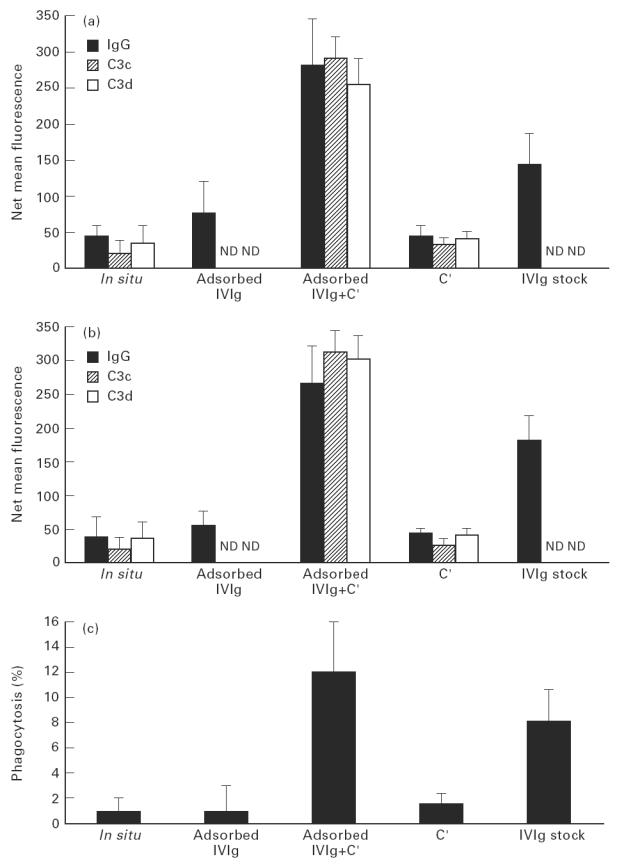
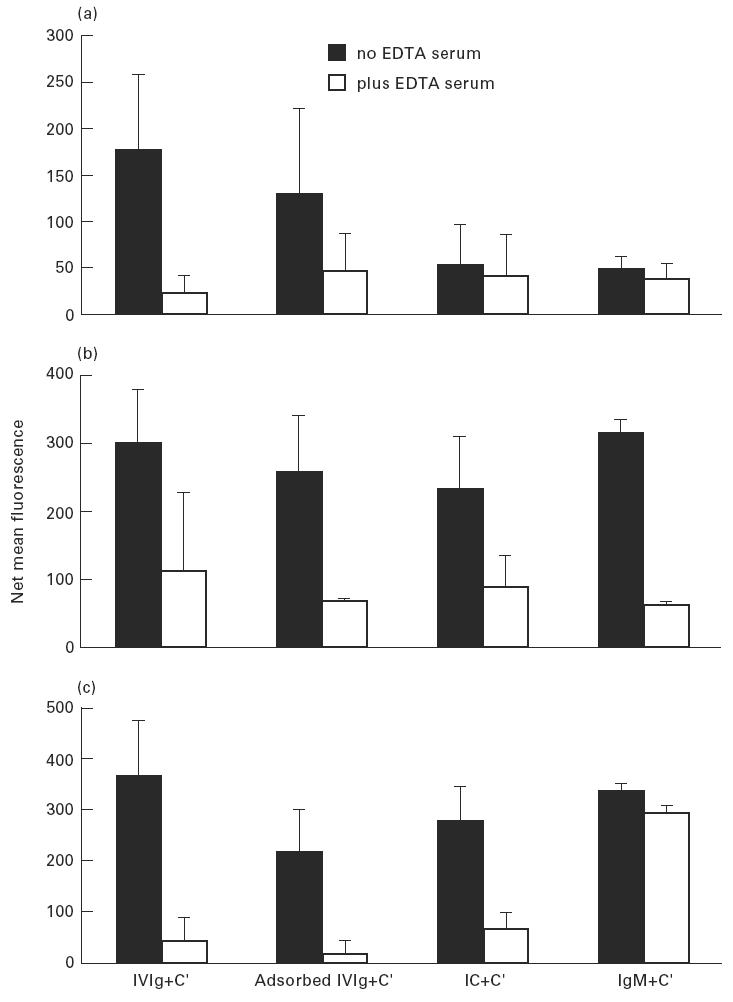
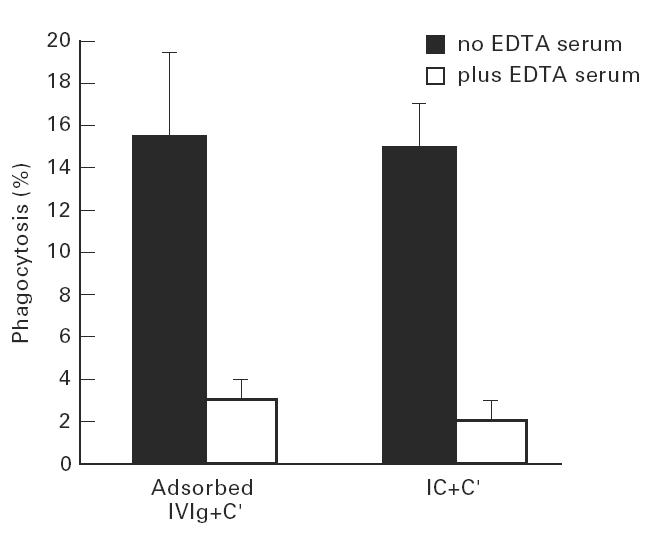
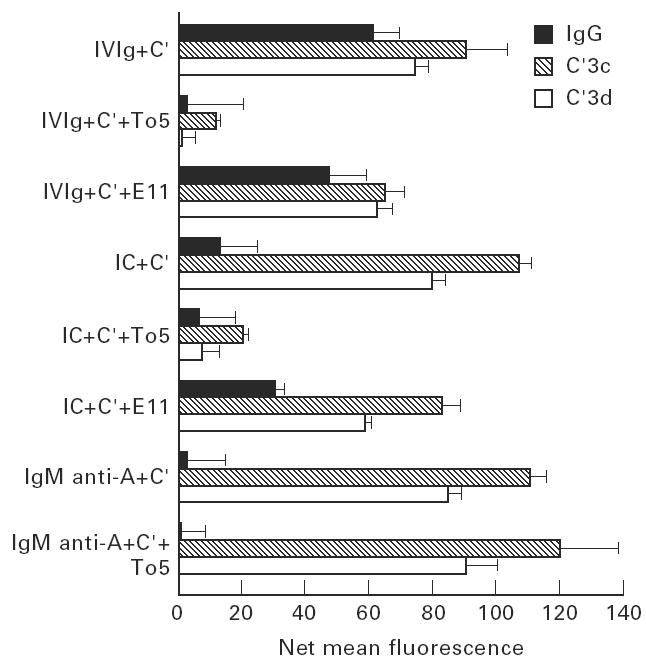
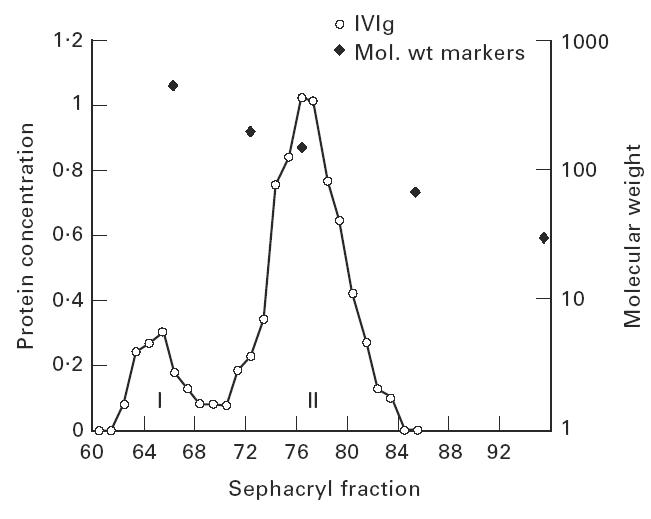
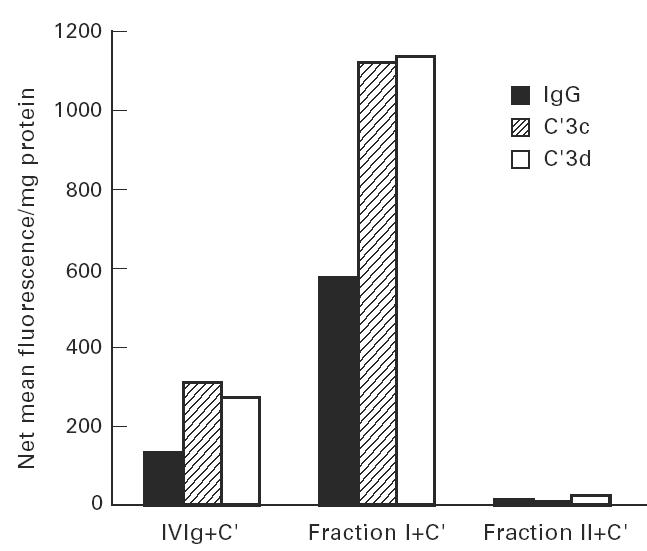
Treatment with i.v.Ig can, on rare occasions, lead to detrimental effects such as enhanced erythrocyte sequestration and an increase in serum immune complexes with inflammatory sequellae such as exacerbation of glomerular nephritis. In this study, i.v.Ig (Sandoglobin) was examined for complement binding moieties which resemble immune complexes and can mediate the binding of IgG and C'3b to human erythrocytes via CR1 and enhance erythrocyte susceptibility to sequestration. Sephacryl S-200 HR separated i.v.Ig into two fractions: monomeric IgG (74%) and larger complexes of the molecular weight of an IgG dimer or greater (> or = 300 kD) (26%). In the presence of complement, the 'dimers' bound to human erythrocytes, rendering them susceptible to phagocytosis in vitro. Removal of erythrocyte-specific isoantibodies from the i.v.Ig had no effect on 'dimer' binding to the erythrocytes. Monomeric IgG contained virtually no complement-activating, erythrocyte-binding activity. Erythrocyte binding of complement-bearing IgG 'dimers' and subsequent phagocytosis resembles the binding of complement-bearing immune complexes to erythrocyte CR1. Exposure to Factor I leads to the release of complement-bearing IgG 'dimers' from erythrocyte CR1 and to the abrogation of erythrophagocytosis. Binding of complement-bearing IgG 'dimers' to the erythrocyte is blocked by To5, a CR1-specific monoclonal antibody.
Figures
Similar articles
-
Exposure to complement-bearing immune complexes enhances the in vitro sequestration of erythrocytes from young but not elderly donors.Clin Exp Immunol. 1993 Feb;91(2):301-7. doi: 10.1111/j.1365-2249.1993.tb05899.x. Clin Exp Immunol. 1993. PMID: 8428395 Free PMC article.
-
In vivo administration of intravenous immunoglobulin (IVIg) can lead to enhanced erythrocyte sequestration.J Autoimmun. 1999 Aug;13(1):129-35. doi: 10.1006/jaut.1999.0302. J Autoimmun. 1999. PMID: 10441177
-
Erythrocyte enhancement of C3b-mediated phagocytosis by human neutrophils in vitro: a combined effect of the erythrocyte complement receptors CR1 and erythrocyte scavengers to reactive oxygen metabolites (ROM).Immunology. 1985 May;55(1):97-103. Immunology. 1985. PMID: 3158597 Free PMC article.
-
The binding of immune complexes by the erythrocyte complement receptor 1 (CR1).Immunopharmacology. 1992 Sep-Oct;24(2):101-6. doi: 10.1016/0162-3109(92)90016-6. Immunopharmacology. 1992. PMID: 1473961 Review.
-
Heteropolymer-mediated clearance of immune complexes via erythrocyte CR1: mechanisms and applications.Immunol Rev. 2001 Oct;183:10-24. doi: 10.1034/j.1600-065x.2001.1830102.x. Immunol Rev. 2001. PMID: 11782244 Review.
Cited by
-
Acute hemolysis after high-dose intravenous immunoglobulin therapy in highly HLA sensitized patients.Clin J Am Soc Nephrol. 2009 Dec;4(12):1993-7. doi: 10.2215/CJN.04540709. Epub 2009 Oct 15. Clin J Am Soc Nephrol. 2009. PMID: 19833910 Free PMC article.
-
On the dark side of therapies with immunoglobulin concentrates: the adverse events.Front Immunol. 2015 Feb 5;6:11. doi: 10.3389/fimmu.2015.00011. eCollection 2015. Front Immunol. 2015. PMID: 25699039 Free PMC article. Review.
References
-
- Mouthon L, Kaveri SV, Spalter SH, et al. Mechanisms of action of intravenous immune globulin in immune-mediated diseases. Clin Exp Immunol. 1996;104:3–9. - PubMed
-
- Miletic VD, Hester CG, Frank MM. Regulation of complement activity by immunoglobulin: effect of immunoglobulin isotype on C4 uptake on antibody-sensitized sheep erythrocytes and solid phase immune complexes. J Immunol. 1996;156:749–57. - PubMed
-
- Basta M. Modulation of complement-mediated immune damage by intravenous immune globulin. Clin Exp Immunol. 1996;104:21–25. - PubMed
-
- Roux KH, Tankersley DL. A view of the human idiotypic repertoire. Electron microscopic and immunologic analyses of spontaneous idiotype–anti-idiotype dimers in pooled human IgG. J Immunol. 1990;144:1387–95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
