

Maturation-dependent vulnerability of oligodendrocytes to oxidative stress-induced death caused by glutathione depletion
- PMID: 9698317
- PMCID: PMC6793198
- DOI: 10.1523/JNEUROSCI.18-16-06241.1998
Maturation-dependent vulnerability of oligodendrocytes to oxidative stress-induced death caused by glutathione depletion
Abstract
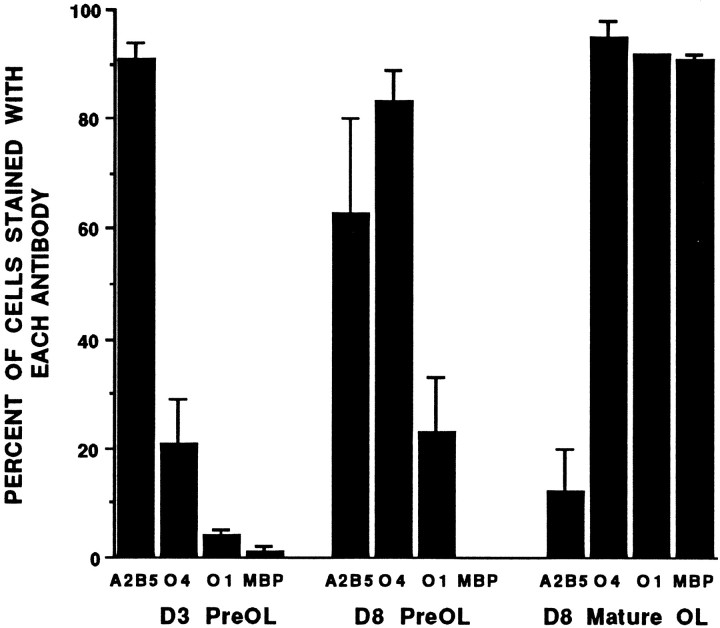
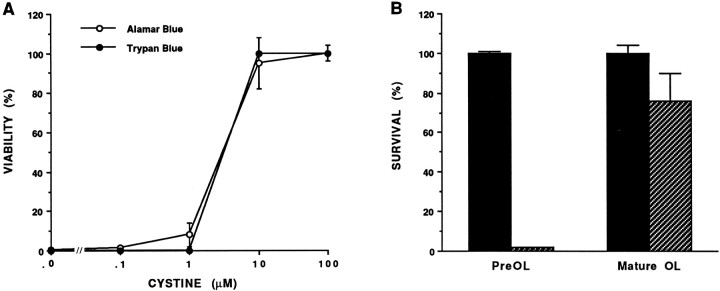
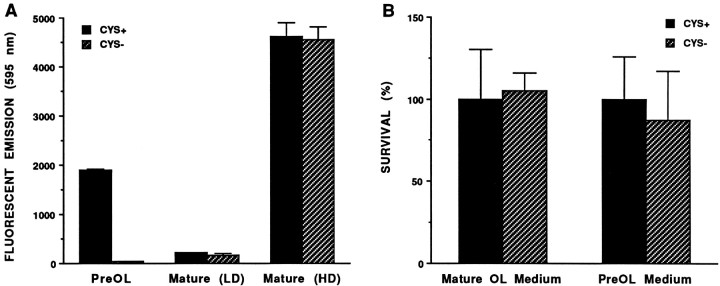
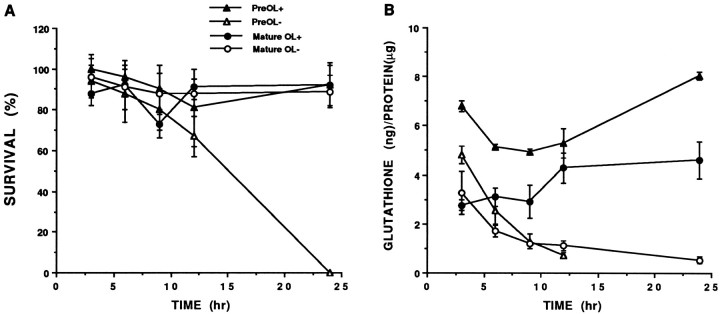
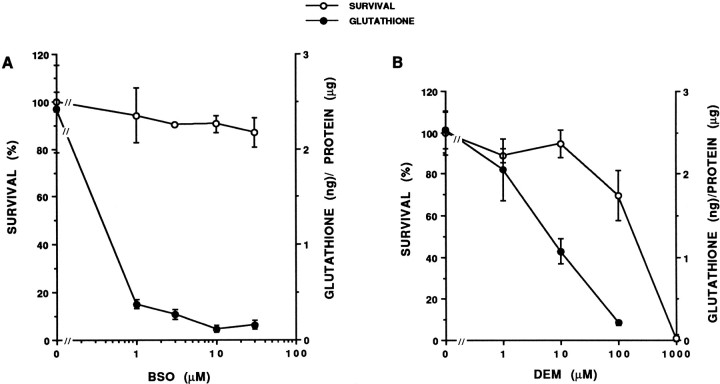
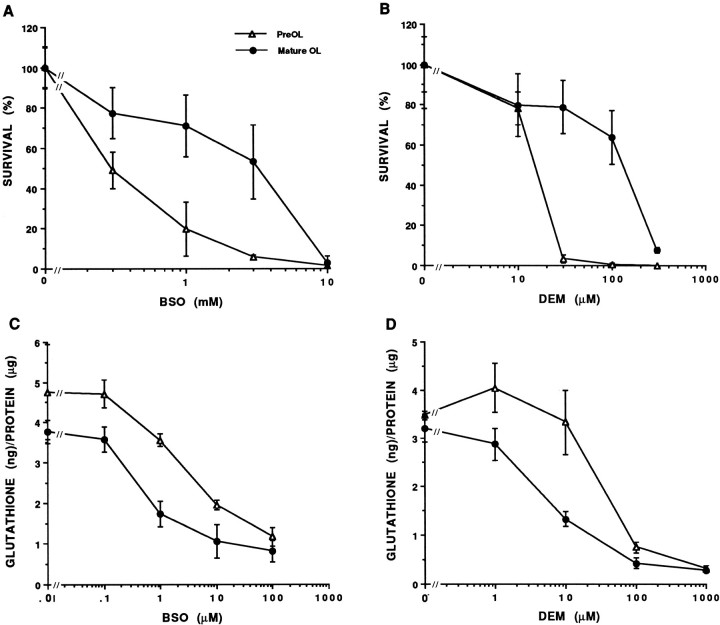
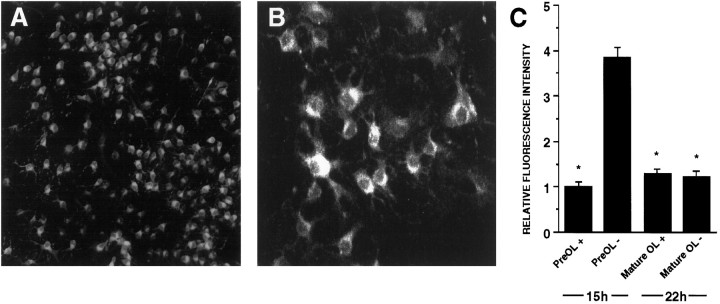
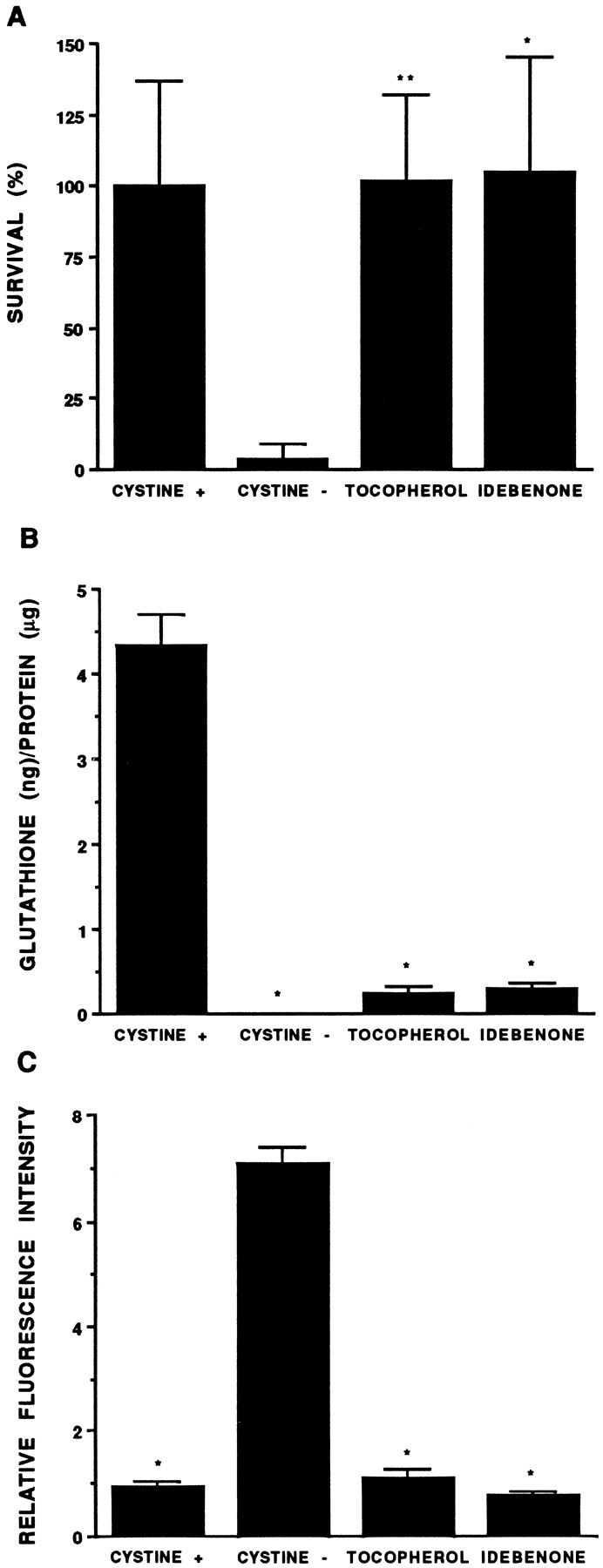
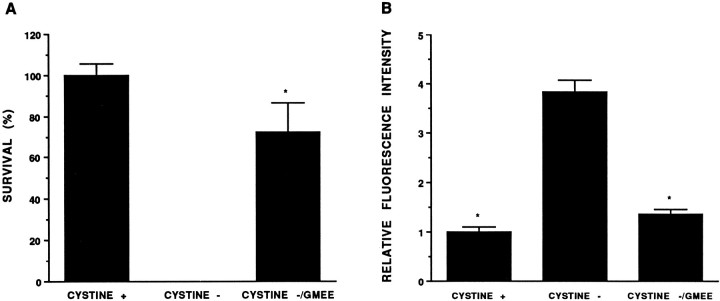
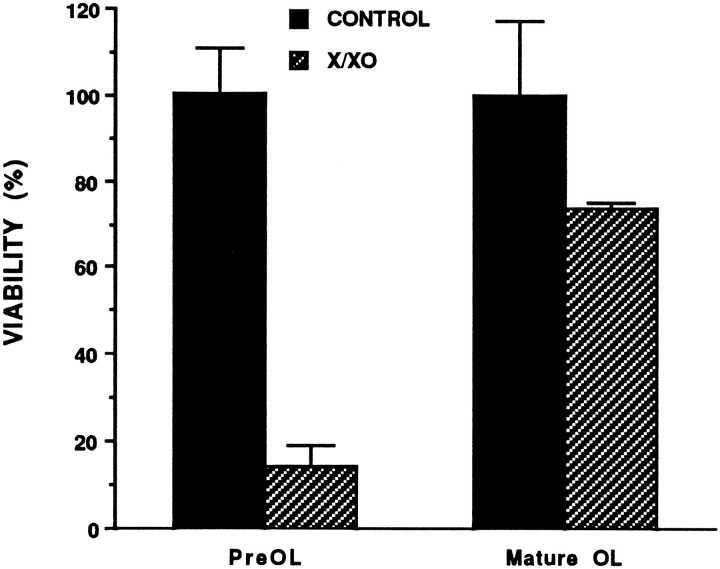
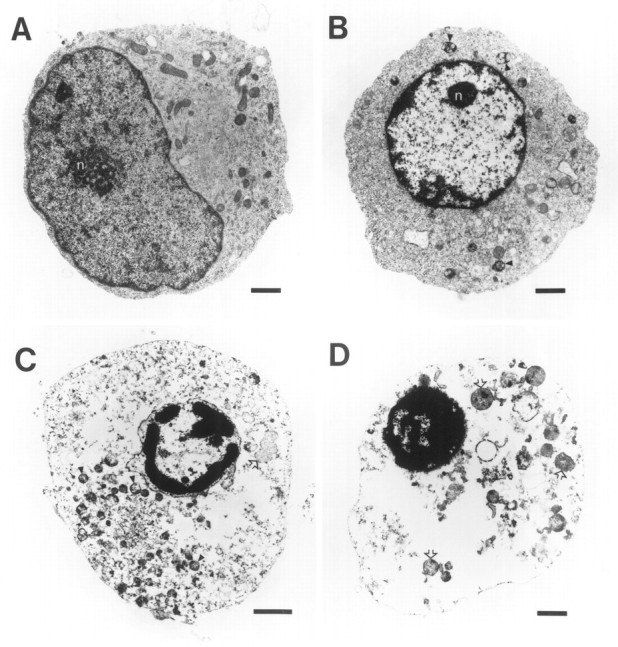
Death of oligodendrocyte (OL) precursors can be triggered in vitro by cystine deprivation, a form of oxidative stress that involves depletion of intracellular glutathione. We report here that OLs demonstrate maturation-dependent differences in survival when subjected to free radical-mediated injury induced by glutathione depletion. Using immunopanning to isolate rat preoligodendrocytes (preOLs), we generated highly enriched populations of preOLs and mature OLs under chemically defined conditions. Cystine deprivation caused a similar decrease in glutathione levels in OLs at both stages. However, preOLs were completely killed by cystine deprivation, whereas mature OLs remained viable. Although the glutathione-depleting agents buthionine sulfoximine and diethylmaleate were more potent in depleting glutathione in mature OLs, both agents were significantly more toxic to preOLs. Glutathione depletion markedly increased intracellular free radical generation in preOLs, but not in mature OLs, as indicated by oxidation of the redox-sensitive probe dihydrorhodamine 123. The antioxidants alpha-tocopherol, idebenone, and glutathione monoethylester prevented the oxidation of dihydrorhodamine in cystine-depleted preOLs and markedly protected against cell death. When the intracellular glutathione level was not manipulated, preOLs were also more vulnerable than mature OLs to exogenous free radical toxicity generated by a xanthine-xanthine oxidase system. Ultrastructural features of free radical-mediated injury in glutathione-depleted preOLs included nuclear condensation, margination of chromatin, and mitochondrial swelling. These observations indicate that preOLs are significantly more sensitive to the toxic effects of glutathione depletion and that oligodendroglial maturation is associated with decreased susceptibility to oxidative stress.
Figures
References
-
- Aizenman E. Modulation of N-methyl-d-aspartate receptors by hydroxyl radicals in rat cortical neurons in vitro. Neurosci Lett. 1995;189:57–59. - PubMed
-
- Anderson ME, Levy EJ, Meister A. Preparation and use of glutathione monoesters. In: Packer L, editor. Oxygen radicals in biological systems, Vol 234, Part D. Academic; San Diego: 1994. pp. 492–499. - PubMed
-
- Back SA, Volpe JJ. Cellular and molecular pathogenesis of periventricular white matter injury. MRDD Research Reviews. 1997;3:96–107.
-
- Back SA, Yonezawa M, Gan X, Rosenberg PA, Volpe JJ. Oligodendrocyte death induced by cystine deprivation occurs by apoptosis. Soc Neurosci Abstr. 1995;21:41.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources