

Potassium current development and its linkage to membrane expansion during growth of cultured embryonic mouse hippocampal neurons: sensitivity to inhibitors of phosphatidylinositol 3-kinase and other protein kinases
- PMID: 9698319
- PMCID: PMC6793179
- DOI: 10.1523/JNEUROSCI.18-16-06261.1998
Potassium current development and its linkage to membrane expansion during growth of cultured embryonic mouse hippocampal neurons: sensitivity to inhibitors of phosphatidylinositol 3-kinase and other protein kinases
Abstract
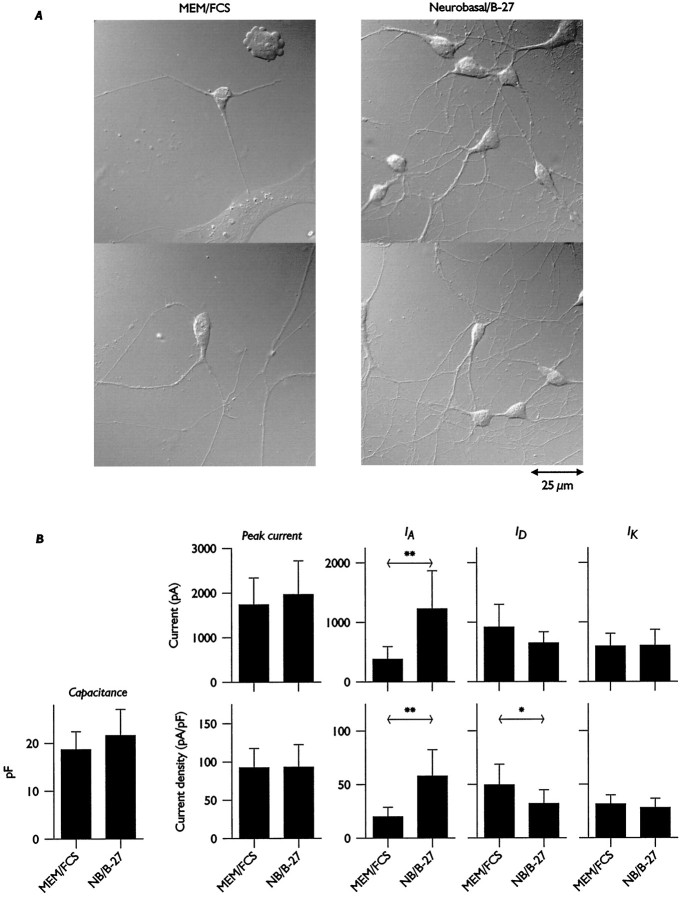
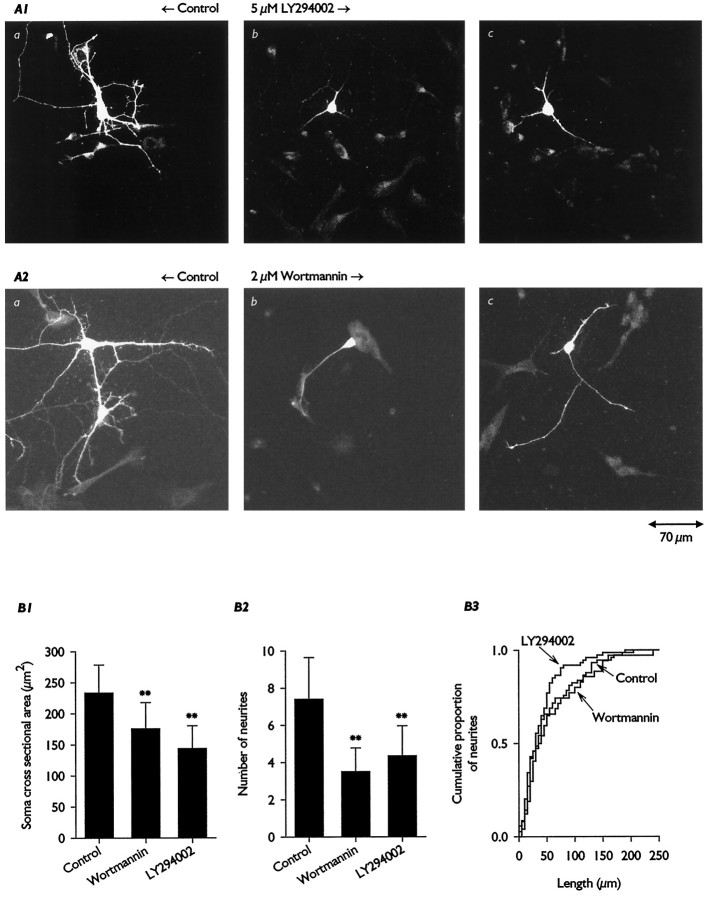
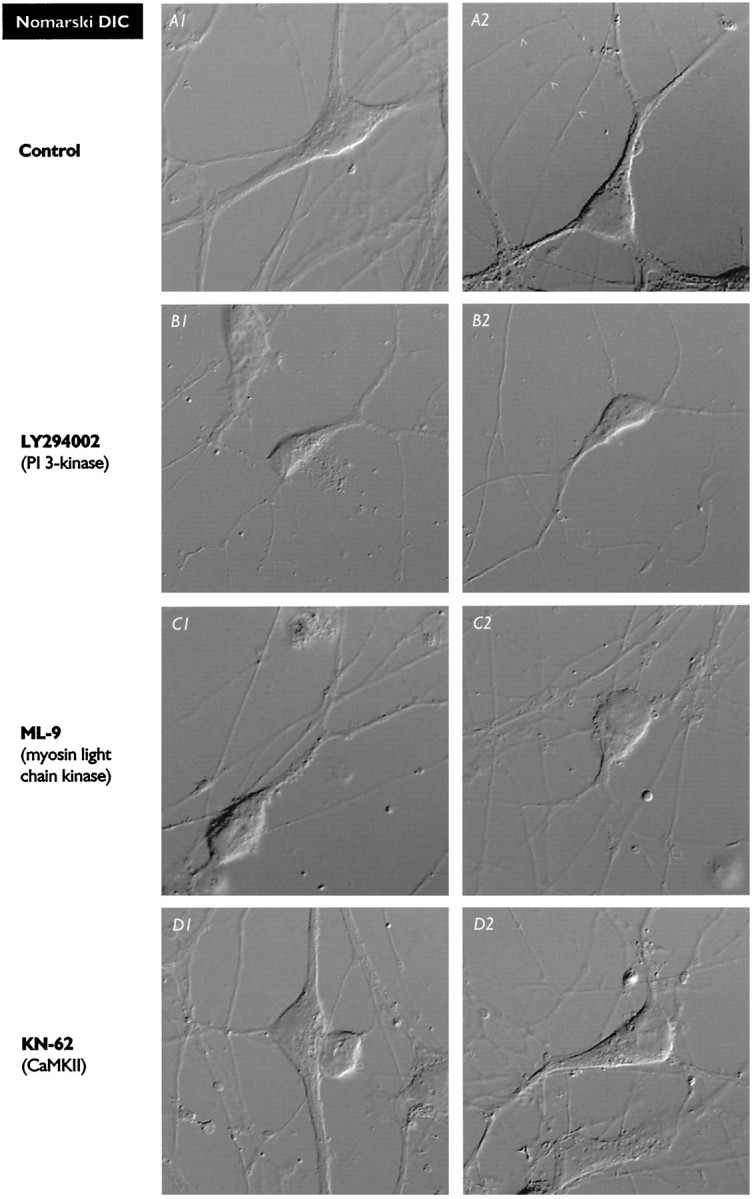
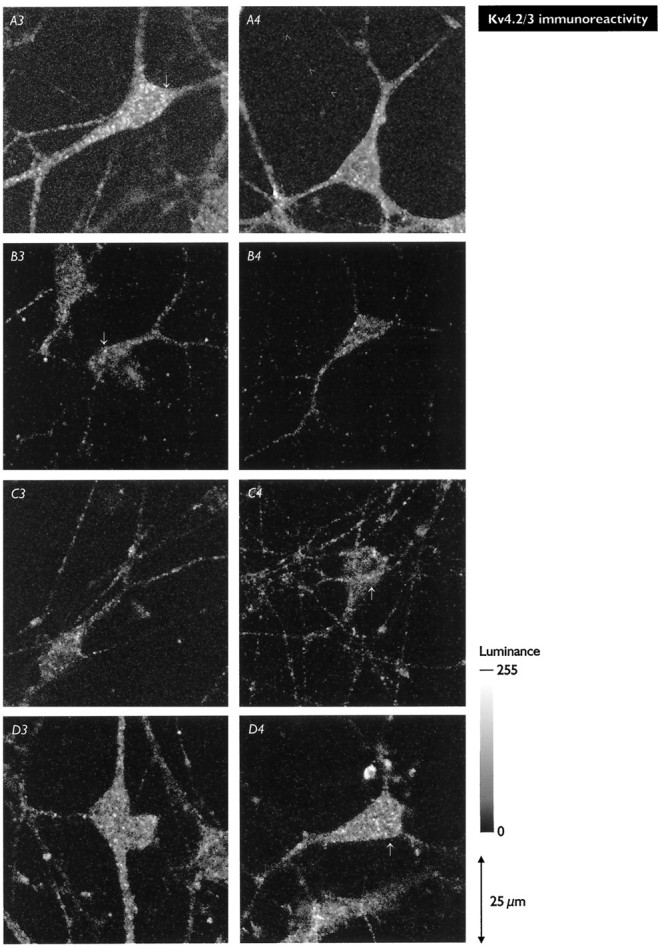
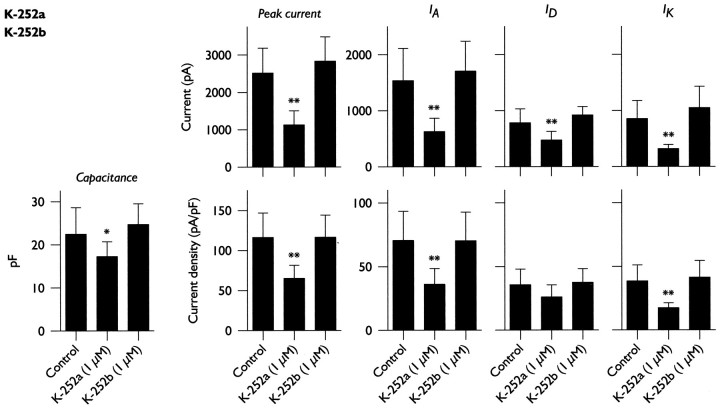
Hippocampal pyramidal neurons express three major voltage-dependent potassium currents, IA, ID, and IK. During hippocampal development, IA, the rapidly activating and inactivating transient potassium current, is detected soon after pyramidal neurons can be morphologically identified. Appearance of IA in developing pyramidal neurons is dependent on contact with cocultured astroglial cells; cultured pyramidal neurons not in contact with astroglial cells have reduced membrane area and IA (Wu and Barish, 1994). We have examined intracellular signaling pathways that could contribute to the regulation of IA development by probing developing pyramidal neurons with kinase inhibitors. We observed that exposure to LY294002 or wortmannin, inhibitors of phosphatidylinositol (PI) 3-kinase, reduced somatic cross-sectional area, neurite outgrowth, whole-cell capacitance, IA amplitude and density (amplitude normalized to membrane area), and immunoreactivity for Kv4.2 and/or Kv4.3 (potassium channel subunits likely to be present in the channels carrying IA). In contrast, exposure to ML-9 or KN-62, inhibitors of myosin light chain kinase or Ca2+-calmodulin-dependent protein kinase II (CaMKII), reduced membrane area and IA amplitude but did not affect IA density or Kv4. 2/3 immunoreactivity to the same extent as inhibitors of PI 3-kinase. Unexpectedly, exposure to bisindolymaleimide I or calphostin C, inhibitors of protein kinase C (PKC), did not affect membrane area or potassium current development. Our data suggest that PI 3-kinases regulate both A-type potassium channel synthesis and plasmalemmal insertion of vesicles bearing these potassium channels. CaMKII appears to regulate fusion of channel-bearing vesicles with the plasmalemma and myosin light chain kinase to regulate centripetal transport of channel-bearing vesicles from the Golgi. We further suggest that astroglial cells exert their influence on pyramidal neuron development through activation of PI 3-kinases.
Figures






Similar articles
-
Regulation of an inactivating potassium current (IA) by the extracellular matrix protein vitronectin in embryonic mouse hippocampal neurones.J Physiol. 2003 Mar 15;547(Pt 3):859-71. doi: 10.1113/jphysiol.2002.036889. Epub 2003 Jan 24. J Physiol. 2003. PMID: 12562917 Free PMC article.
-
Astroglial modulation of transient potassium current development in cultured mouse hippocampal neurons.J Neurosci. 1994 Mar;14(3 Pt 2):1677-87. doi: 10.1523/JNEUROSCI.14-03-01677.1994. J Neurosci. 1994. PMID: 7510334 Free PMC article.
-
Calcium-calmodulin-dependent kinase II modulates Kv4.2 channel expression and upregulates neuronal A-type potassium currents.J Neurosci. 2004 Apr 7;24(14):3643-54. doi: 10.1523/JNEUROSCI.0154-04.2004. J Neurosci. 2004. PMID: 15071113 Free PMC article.
-
Two pharmacologically and kinetically distinct transient potassium currents in cultured embryonic mouse hippocampal neurons.J Neurosci. 1992 Jun;12(6):2235-46. doi: 10.1523/JNEUROSCI.12-06-02235.1992. J Neurosci. 1992. PMID: 1607938 Free PMC article.
-
Voltage-dependent currents of vertebrate neurons and their role in membrane excitability.Adv Neurol. 1986;44:137-70. Adv Neurol. 1986. PMID: 2422889 Review.
Cited by
-
Human induced pluripotent stem cell-derived models to investigate human cytomegalovirus infection in neural cells.PLoS One. 2012;7(11):e49700. doi: 10.1371/journal.pone.0049700. Epub 2012 Nov 27. PLoS One. 2012. PMID: 23209593 Free PMC article.
-
Modulation of a slowly inactivating potassium current, I(D), by metabotropic glutamate receptor activation in cultured hippocampal pyramidal neurons.J Neurosci. 1999 Aug 15;19(16):6825-37. doi: 10.1523/JNEUROSCI.19-16-06825.1999. J Neurosci. 1999. PMID: 10436040 Free PMC article.
-
Somato-dendritic decoupling as a novel mechanism for protracted cortical maturation.BMC Biol. 2016 Jun 21;14:48. doi: 10.1186/s12915-016-0270-5. BMC Biol. 2016. PMID: 27328836 Free PMC article.
-
Interleukin 33-mediated inhibition of A-type K+ channels induces sensory neuronal hyperexcitability and nociceptive behaviors in mice.Theranostics. 2022 Feb 14;12(5):2232-2247. doi: 10.7150/thno.69320. eCollection 2022. Theranostics. 2022. PMID: 35265208 Free PMC article.
-
Regulation of an inactivating potassium current (IA) by the extracellular matrix protein vitronectin in embryonic mouse hippocampal neurones.J Physiol. 2003 Mar 15;547(Pt 3):859-71. doi: 10.1113/jphysiol.2002.036889. Epub 2003 Jan 24. J Physiol. 2003. PMID: 12562917 Free PMC article.
References
-
- Audesirk G, Cablee L, Kern M. Modulation of neurite branching by protein phosphorylation in cultured rat hippocampal neurons. Dev Brain Res. 1997;102:247–260. - PubMed
-
- Banker GA, Cowan WM. Rat hippocampal neurons in dispersed cell culture. Brain Res. 1977;126:397–425. - PubMed
-
- Banker GA, Cowan WM. Further observations on hippocampal neurons in dispersed cell culture. J Comp Neurol. 1979;187:469–494. - PubMed
-
- Barish ME. Modulation of the electrical differentiation of neurons by interactions with glia and other non-neuronal cells. Perspect Dev Neurobiol. 1995;2:357–370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous