

Brain endothelial cells express cyclooxygenase-2 during lipopolysaccharide-induced fever: light and electron microscopic immunocytochemical studies
- PMID: 9698320
- PMCID: PMC6793199
- DOI: 10.1523/JNEUROSCI.18-16-06279.1998
Brain endothelial cells express cyclooxygenase-2 during lipopolysaccharide-induced fever: light and electron microscopic immunocytochemical studies
Abstract
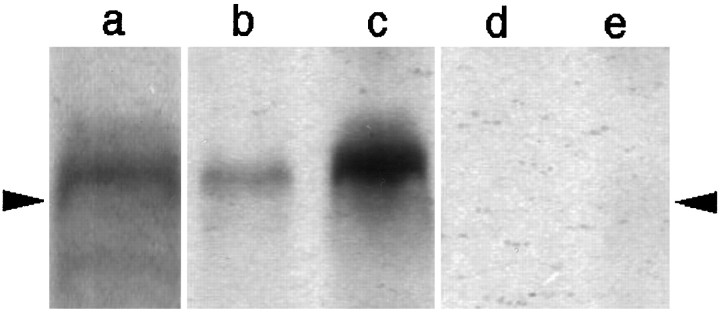
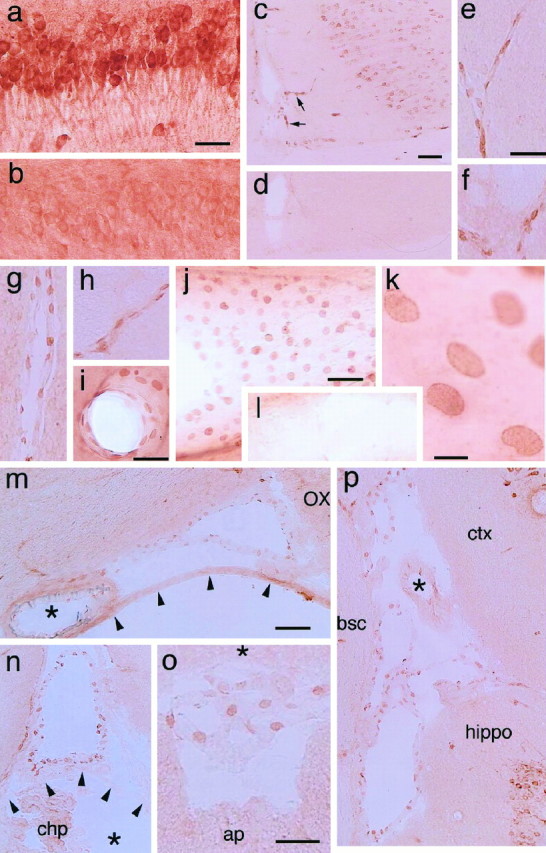
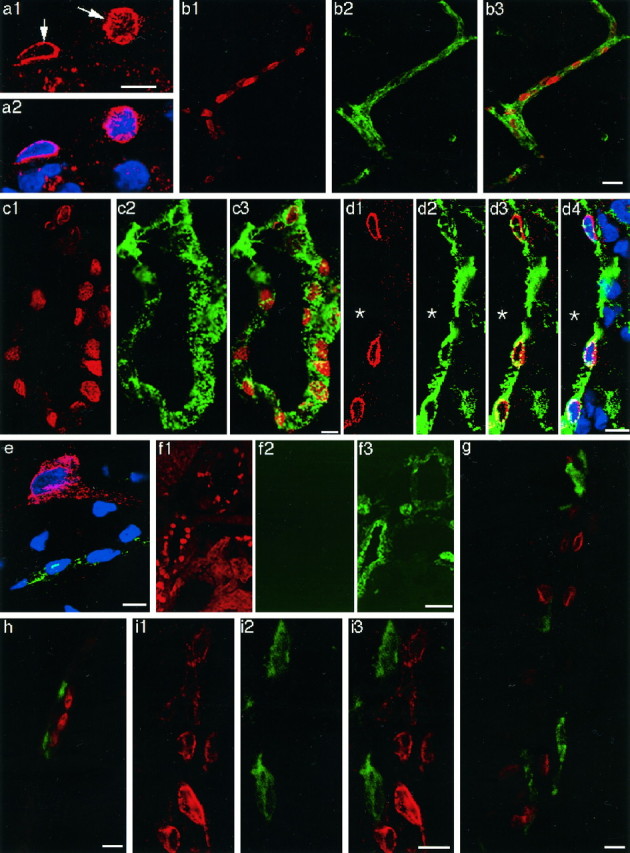
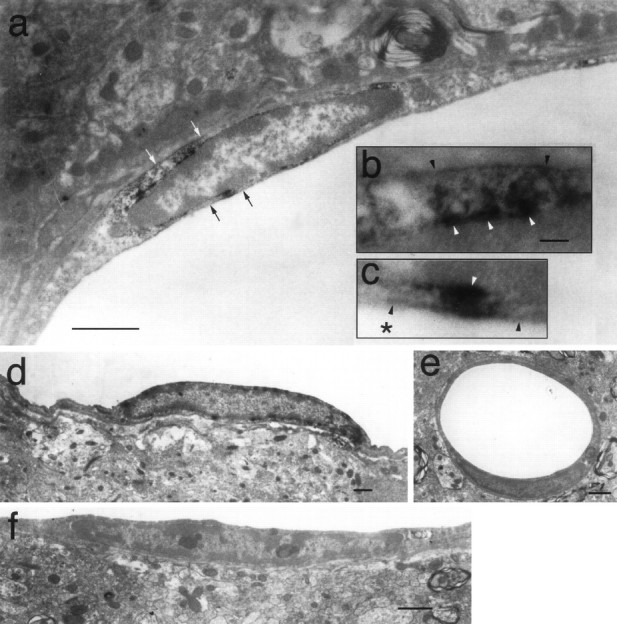
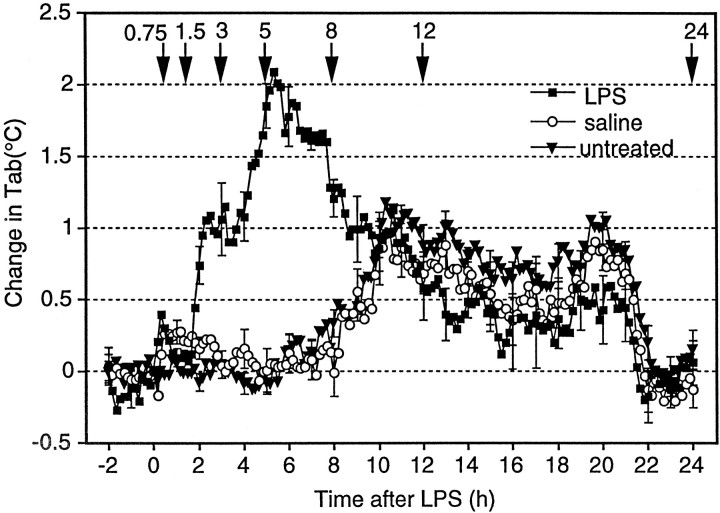
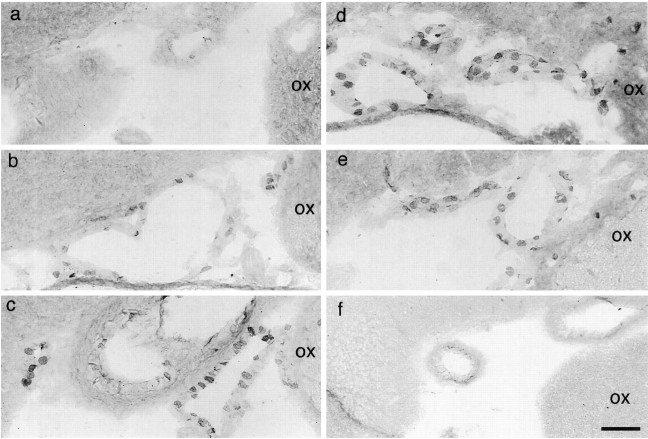
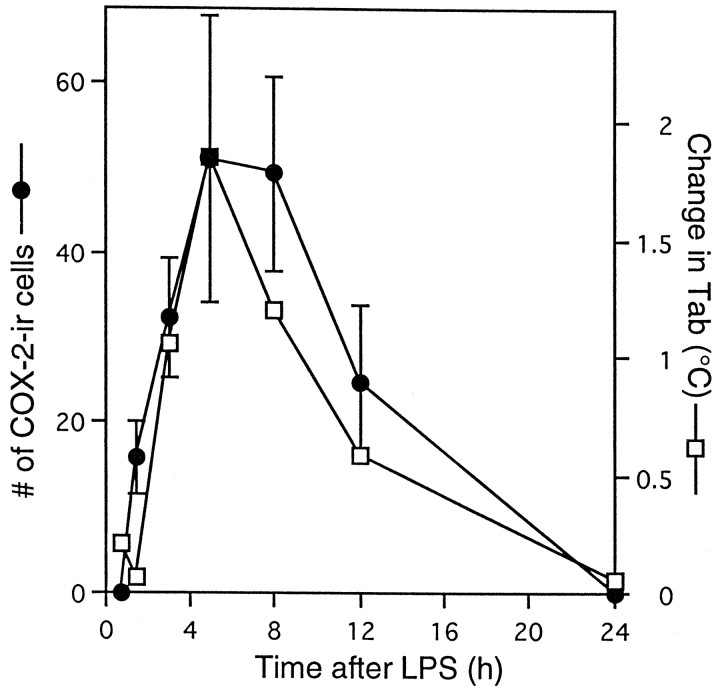
Cyclooxygenase-2 (COX-2), a key enzyme in the biosynthesis of prostaglandins, is induced in brain blood vessels by pyrogens, and its essential role in fever has been hypothesized. In this study, we determined (1) the type of cells that express cyclooxygenase-2 in brain blood vessels of lipopolysaccharide-treated rats, and (2) the precise relationship between the time course of fever and that of cyclooxygenase-2 protein expression in these cells. Five hours after the lipopolysaccharide injection (100 microg/kg, i.p.), cyclooxygenase-2-like immunoreactive cells were found in the parenchymal and subarachnoidal blood vessels. In these blood vessels, the cyclooxygenase-2-like immunoreactivity was restricted to the perinuclear region of the endothelial cells as revealed by a laser confocal microscopy, double-immunofluorescence staining with an endothelial marker, and immunoelectron microscopy. On the other hand, the cyclooxygenase-2-like immunoreactive cells were distinct from microglia or perivascular/meningeal macrophages as revealed by double immunostaining with macrophage/microglia-specific antibodies. Cyclooxygenase-2-like immunoreactive cells were first found at 1.5 hr after the lipopolysaccharide injection, at which time the fever had not been developed. After that, the number of cyclooxygenase-2-like immunoreactive cells and fever followed a similar time course, both being highest at 5 hr after the lipopolysaccharide injection and both returning to the baseline by 24 hr. These results demonstrate that brain endothelial cells are the primary sites where the activation of arachidonic acid cascade takes place during fever after intraperitoneal injection of lipopolysaccharide.
Figures
References
-
- Blalock JE. A molecular basis for bidirectional communication between the immune and neuroendocrine systems. Physiol Rev. 1989;69:1–32. - PubMed
-
- Blatteis CM, Sehic E. Fever: how may circulating pyrogens signal the brain? News Physiol Sci. 1997;12:1–9.
-
- Breder CD, Saper CB. Expression of inducible cyclooxygenase mRNA in the mouse brain after systemic administration of bacterial lipopolysaccharide. Brain Res. 1996;713:64–69. - PubMed
-
- Cao C, Matsumura K, Yamagata K, Watanabe Y. Induction by lipopolysaccharide of cyclooxygenase-2 mRNA in rat brain; its possible role in the febrile response. Brain Res. 1995;697:187–196. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials